

Regulation of the ald gene encoding alanine dehydrogenase by AldR in Mycobacterium smegmatis
- PMID: 23749971
- PMCID: PMC3754569
- DOI: 10.1128/JB.00482-13
Regulation of the ald gene encoding alanine dehydrogenase by AldR in Mycobacterium smegmatis
Abstract
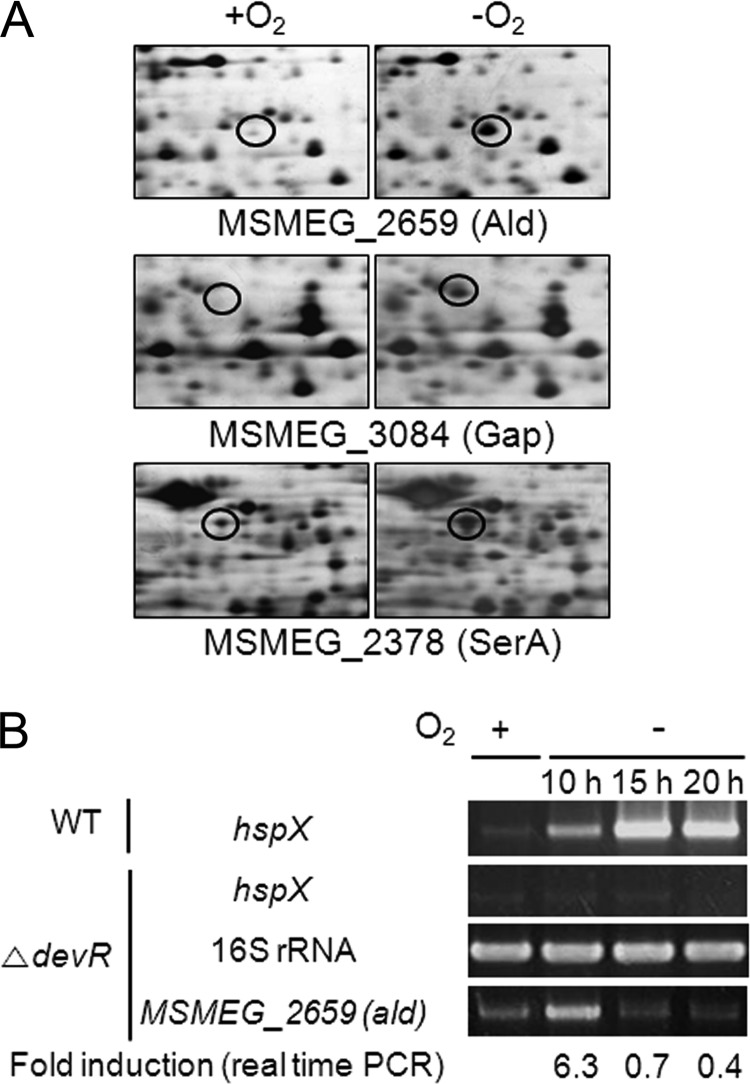
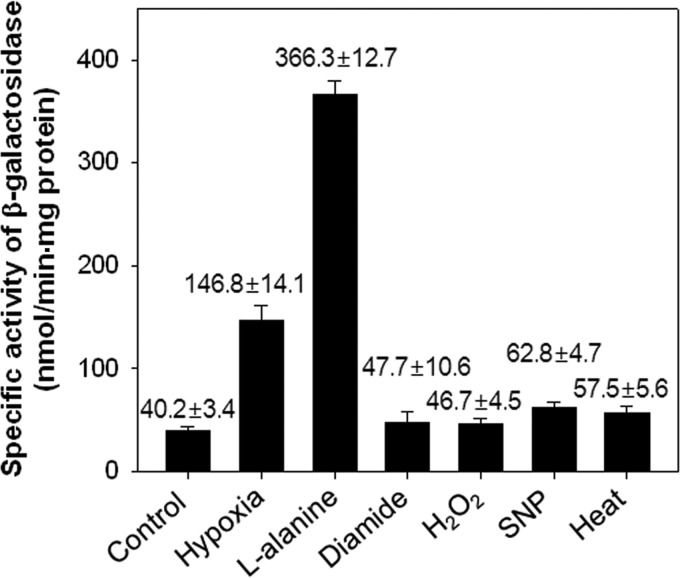
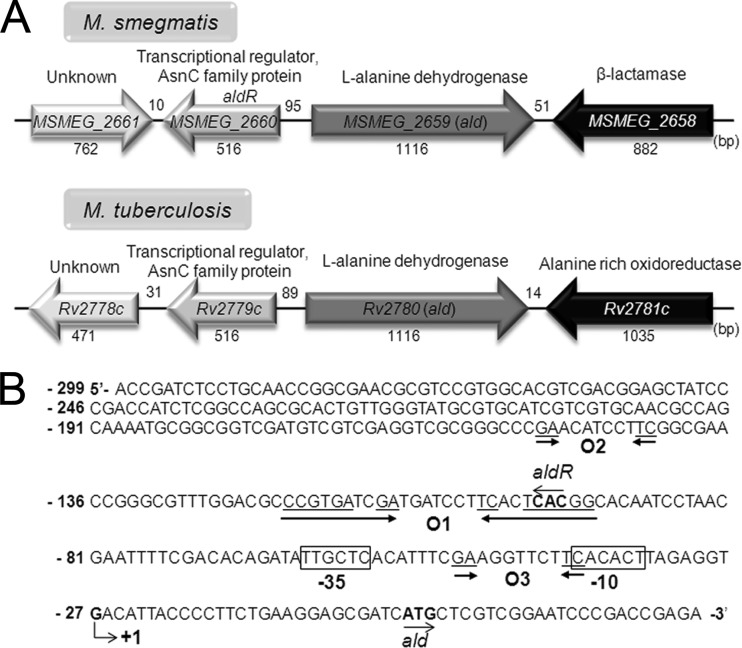
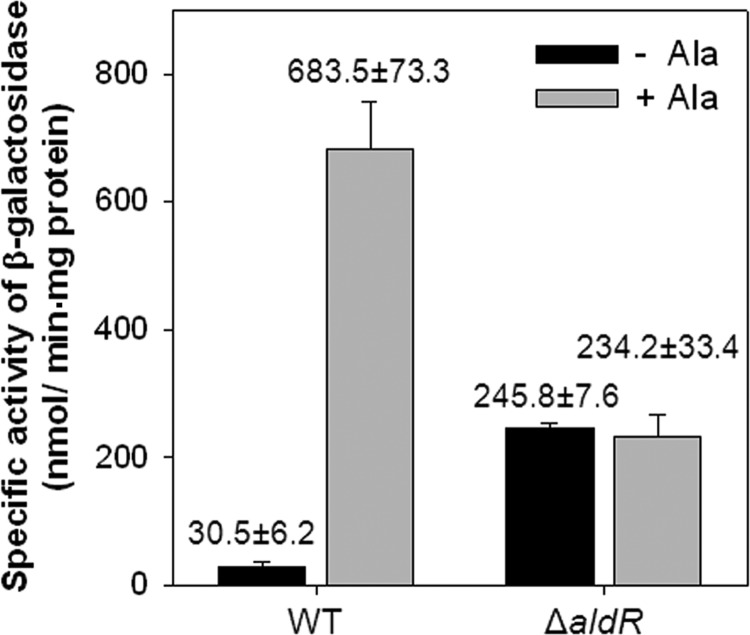
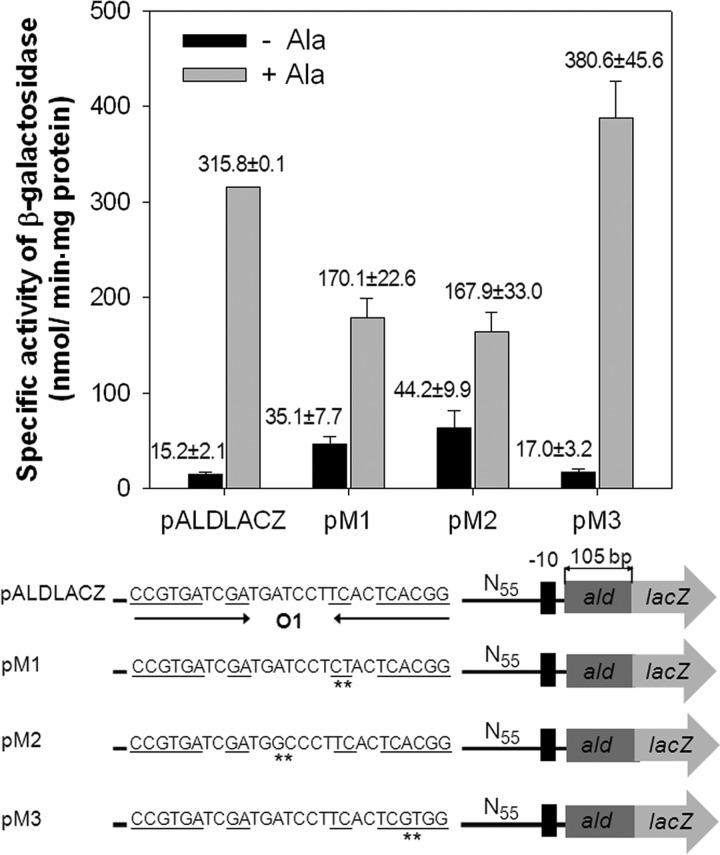
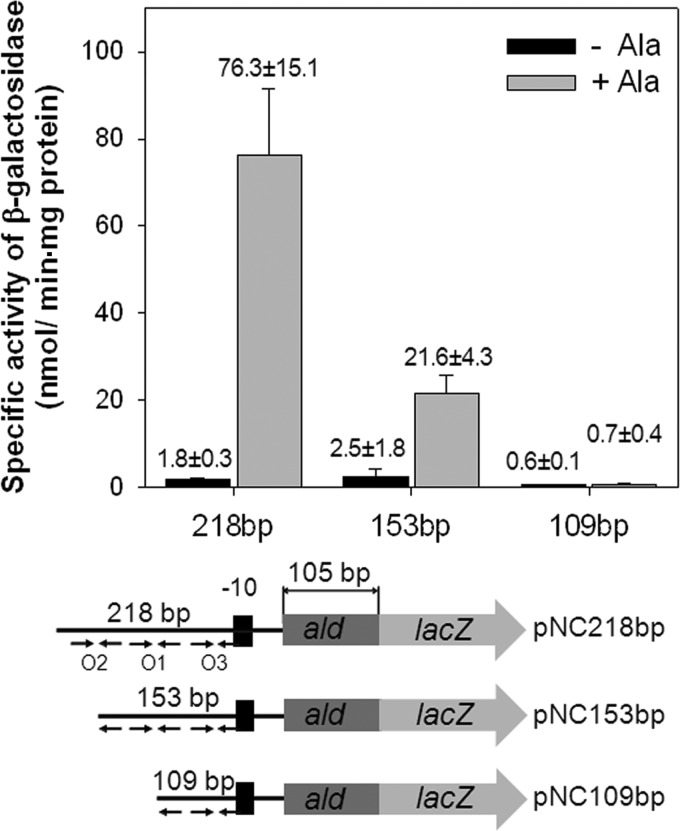
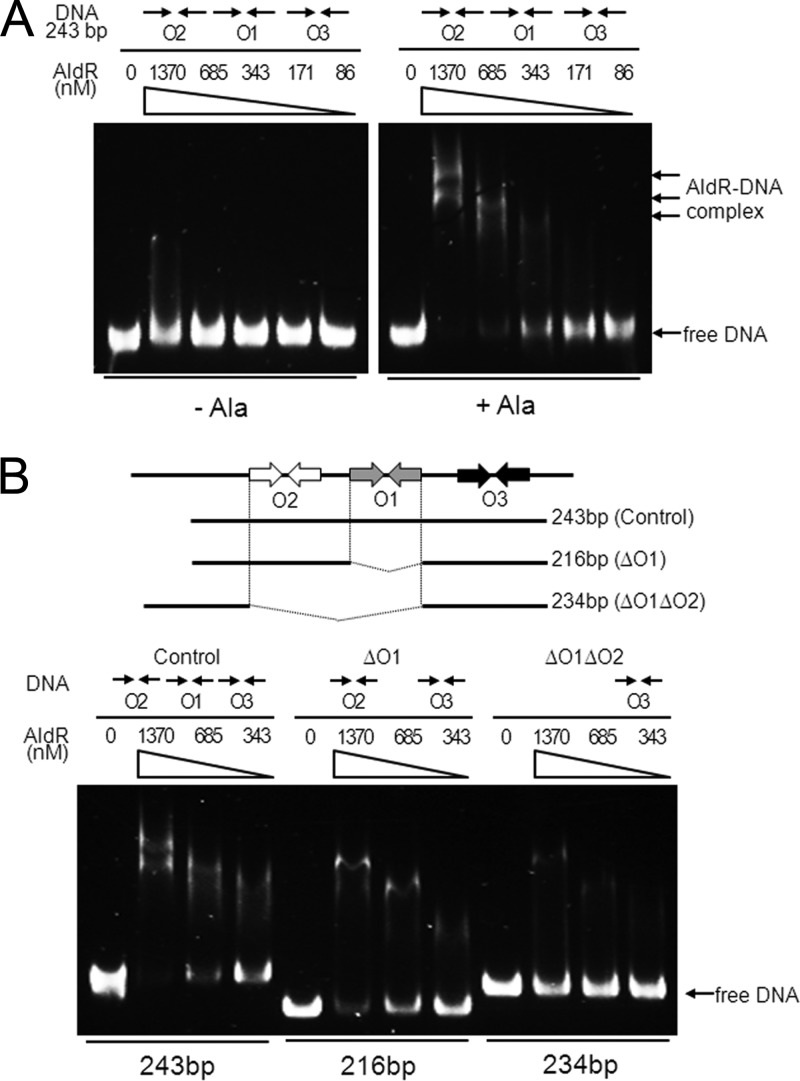
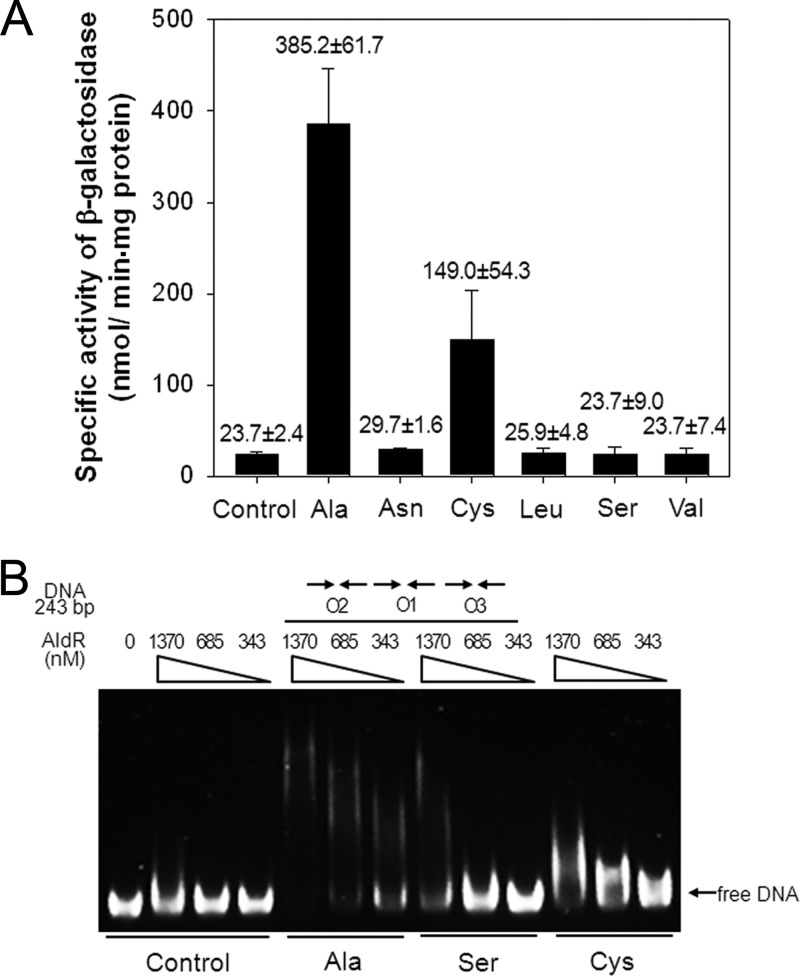
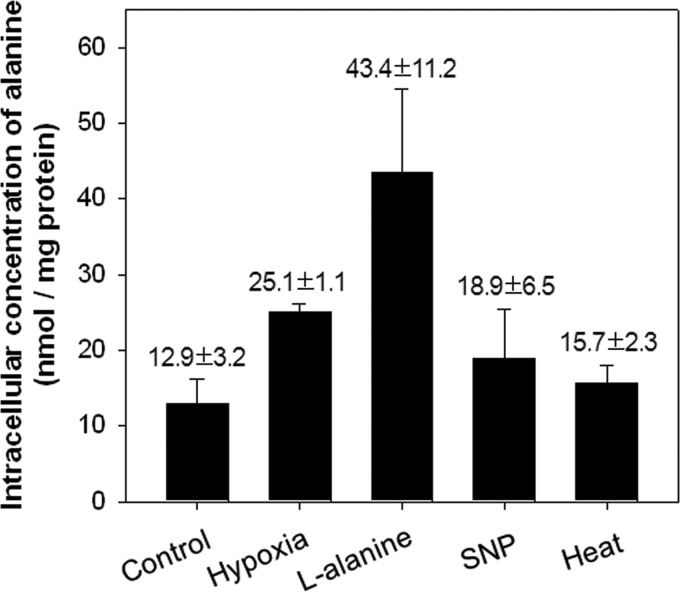
The regulatory gene aldR was identified 95 bp upstream of the ald gene encoding L-alanine dehydrogenase in Mycobacterium smegmatis. The AldR protein shows sequence similarity to the regulatory proteins of the Lrp/AsnC family. Using an aldR deletion mutant, we demonstrated that AldR serves as both activator and repressor for the regulation of ald gene expression, depending on the presence or absence of L-alanine. The purified AldR protein exists as a homodimer in the absence of L-alanine, while it adopts the quaternary structure of a homohexamer in the presence of L-alanine. The binding affinity of AldR for the ald control region was shown to be increased significantly by L-alanine. Two AldR binding sites (O1 and O2) with the consensus sequence GA-N₂-ATC-N₂-TC and one putative AldR binding site with the sequence GA-N₂-GTT-N₂-TC were identified upstream of the ald gene. Alanine and cysteine were demonstrated to be the effector molecules directly involved in the induction of ald expression. The cellular level of L-alanine was shown to be increased in M. smegmatis cells grown under hypoxic conditions, and the hypoxic induction of ald expression appears to be mediated by AldR, which senses the intracellular level of alanine.
Figures
References
-
- Hutter B, Dick T. 1998. Increased alanine dehydrogenase activity during dormancy in Mycobacterium smegmatis. FEMS Microbiol. Lett. 167: 7– 11 - PubMed
-
- Betts JC, Lukey PT, Robb LC, McAdam RA, Duncan K. 2002. Evaluation of a nutrient starvation model of Mycobacterium tuberculosis persistence by gene and protein expression profiling. Mol. Microbiol. 43: 717– 731 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
