

Bioenergetics in human evolution and disease: implications for the origins of biological complexity and the missing genetic variation of common diseases
- PMID: 23754818
- PMCID: PMC3685467
- DOI: 10.1098/rstb.2012.0267
Bioenergetics in human evolution and disease: implications for the origins of biological complexity and the missing genetic variation of common diseases
Abstract
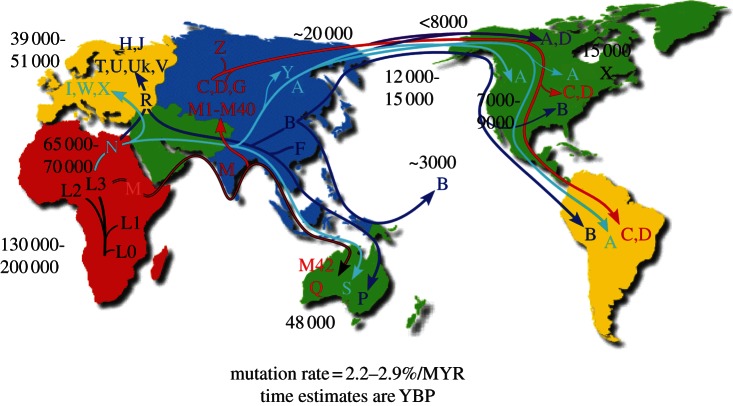
Two major inconsistencies exist in the current neo-Darwinian evolutionary theory that random chromosomal mutations acted on by natural selection generate new species. First, natural selection does not require the evolution of ever increasing complexity, yet this is the hallmark of biology. Second, human chromosomal DNA sequence variation is predominantly either neutral or deleterious and is insufficient to provide the variation required for speciation or for predilection to common diseases. Complexity is explained by the continuous flow of energy through the biosphere that drives the accumulation of nucleic acids and information. Information then encodes complex forms. In animals, energy flow is primarily mediated by mitochondria whose maternally inherited mitochondrial DNA (mtDNA) codes for key genes for energy metabolism. In mammals, the mtDNA has a very high mutation rate, but the deleterious mutations are removed by an ovarian selection system. Hence, new mutations that subtly alter energy metabolism are continuously introduced into the species, permitting adaptation to regional differences in energy environments. Therefore, the most phenotypically significant gene variants arise in the mtDNA, are regional, and permit animals to occupy peripheral energy environments where rarer nuclear DNA (nDNA) variants can accumulate, leading to speciation. The neutralist-selectionist debate is then a consequence of mammals having two different evolutionary strategies: a fast mtDNA strategy for intra-specific radiation and a slow nDNA strategy for speciation. Furthermore, the missing genetic variation for common human diseases is primarily mtDNA variation plus regional nDNA variants, both of which have been missed by large, inter-population association studies.
Keywords: bioenergetics; evolution; mitochondria.
Figures
References
-
- Darwin C. 1859. On the origin of species by means of natural selection, or the preservation of favoured races in the struggle for life. London, UK: John Murray of Albemarle Street
-
- Darwin CR, Wallace AR. 1858. On the tendency of species to form varieties; and on the perpetuation of varieties and species by natural means of selection. J. Proc. Linn. Soc. Lond. Zool. 3, 46–50
-
- Schrodinger E. 1944. What is life? The physical aspect of the living cell. Cambridge, UK: Cambridge University Press
-
- Morowitz HJ. 1968. Energy flow in biology: biological organization as a problem in thermal physics. New York, NY: Academic Press
-
- Lane N. 2011. Energetics and genetics across the prokaryote–eukaryote divide. Biol. Direct 6, 35. 10.1186/1745-6150-6-35 (doi:10.1186/1745-6150-6-35) - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases