

Juvenile hormone and insulin regulate trehalose homeostasis in the red flour beetle, Tribolium castaneum
- PMID: 23754959
- PMCID: PMC3675034
- DOI: 10.1371/journal.pgen.1003535
Juvenile hormone and insulin regulate trehalose homeostasis in the red flour beetle, Tribolium castaneum
Abstract
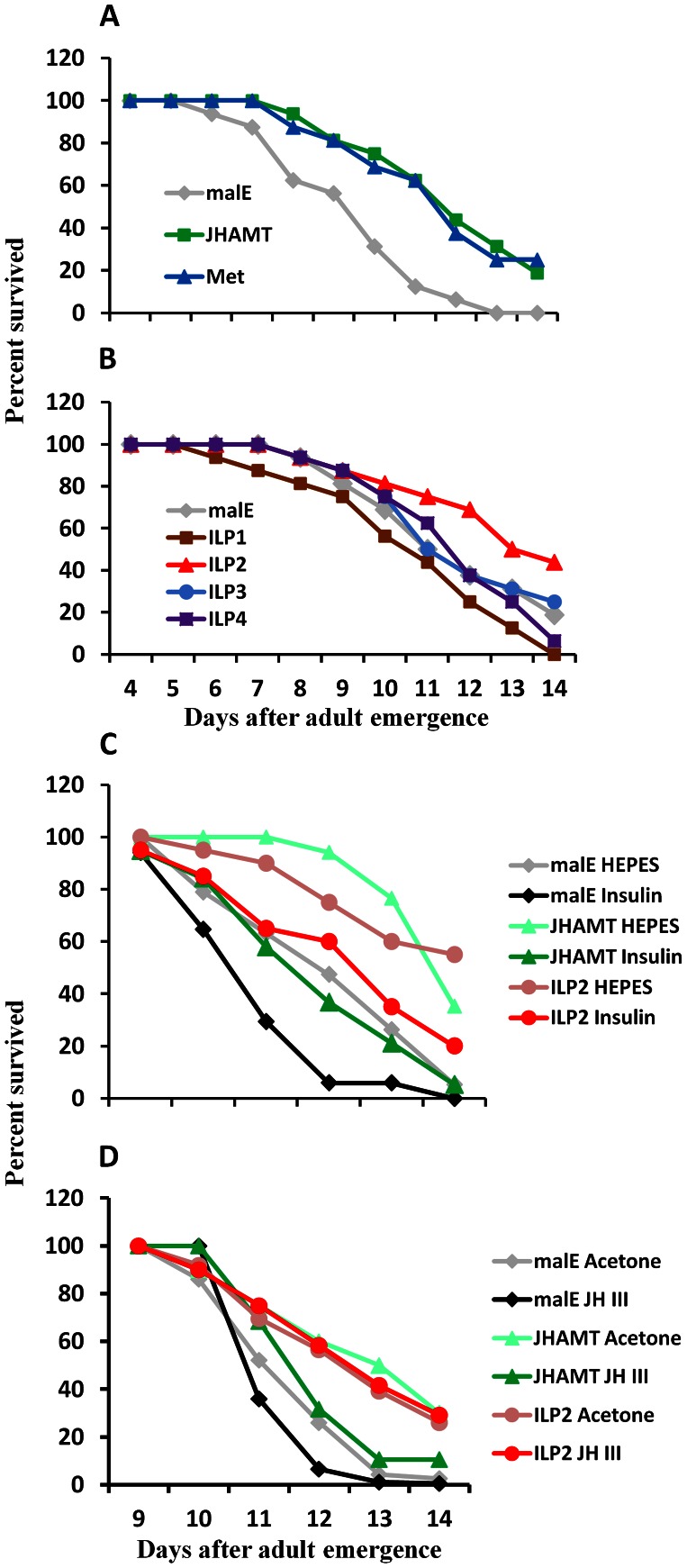
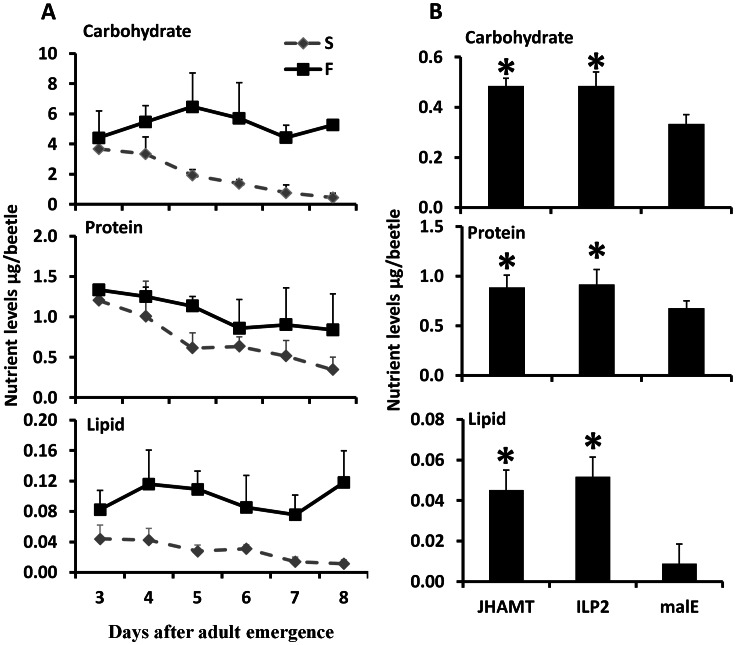
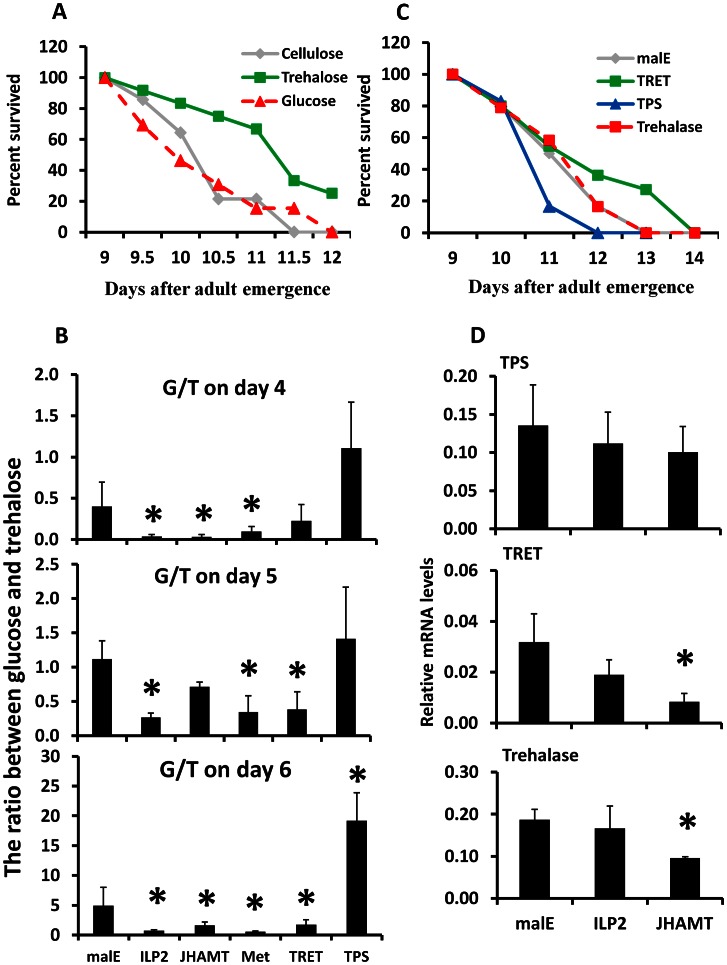
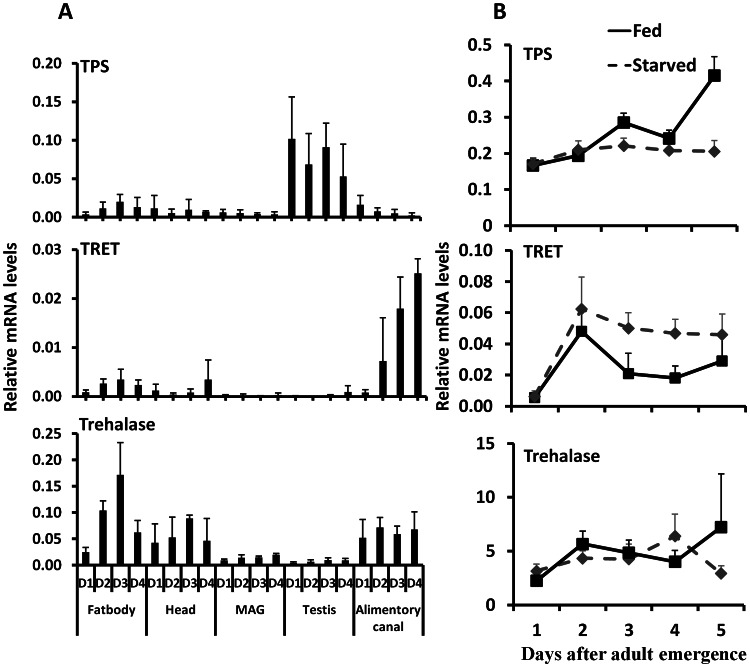
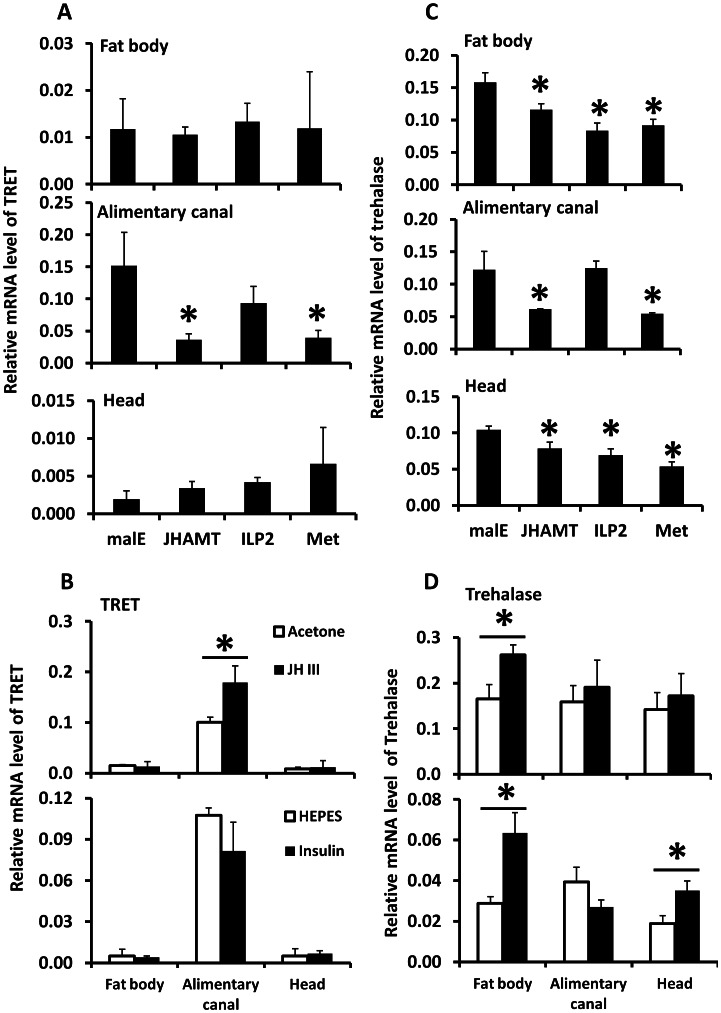
Insulin/IGF-1 signaling (IIS) has been well studied for its role in the control of life span extension and resistance to a variety of stresses. The Drosophila melanogaster insulin-like receptor (InR) mutant showed extended life span due to reduced juvenile hormone (JH) levels. However, little is known about the mechanism of cross talk between IIS and JH in regulation of life span extension and resistance to starvation. In the current study, we investigated the role of IIS and JH signaling in regulation of resistance to starvation. Reduction in JH biosynthesis, JH action, or insulin-like peptide 2 (ILP2) syntheses by RNA interference (RNAi)-aided knockdown in the expression of genes coding for juvenile hormone acid methyltransferase (JHAMT), methoprene-tolerant (Met), or ILP2 respectively decreased lipid and carbohydrate metabolism and extended the survival of starved beetles. Interestingly, the extension of life span could be restored by injection of bovine insulin into JHAMT RNAi beetles but not by application of JH III to ILP2 RNAi beetles. These data suggest that JH controls starvation resistance by regulating synthesis of ILP2. More importantly, JH regulates trehalose homeostasis, including trehalose transport and metabolism, and controls utilization of stored nutrients in starved adults.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Jindra M, Palli SR, Riddiford LM (2013) The Juvenile hormone signaling pathway in insect development. Annu Rev Entomol 58: 181–204. - PubMed
-
- Riddiford LM (2012) How does juvenile hormone control insect metamorphosis and reproduction? Gen Comp Endocrinol 179: 477–484. - PubMed
-
- Minakuchi C, Namiki T, Shinoda T (2009) Kruppel homolog 1, an early juvenile hormone-response gene downstream of Methoprene-tolerant, mediates its anti-metamorphic action in the red flour beetle Tribolium castaneum . Dev Biol 325: 341–350. - PubMed
-
- Truman JW, Hiruma K, Allee JP, Macwhinnie SG, Champlin DT, et al. (2006) Juvenile hormone is required to couple imaginal disc formation with nutrition in insects. Science 312: 1385–1388. - PubMed
-
- Koyama T, Syropyatova MO, Riddiford LM (2008) Insulin/IGF signaling regulates the change in commitment in imaginal discs and primordia by overriding the effect of juvenile hormone. Dev Biol 324: 258–265. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
