

Drug-induced conformational and dynamical changes of the S31N mutant of the influenza M2 proton channel investigated by solid-state NMR
- PMID: 23758317
- PMCID: PMC4004037
- DOI: 10.1021/ja4041412
Drug-induced conformational and dynamical changes of the S31N mutant of the influenza M2 proton channel investigated by solid-state NMR
Abstract
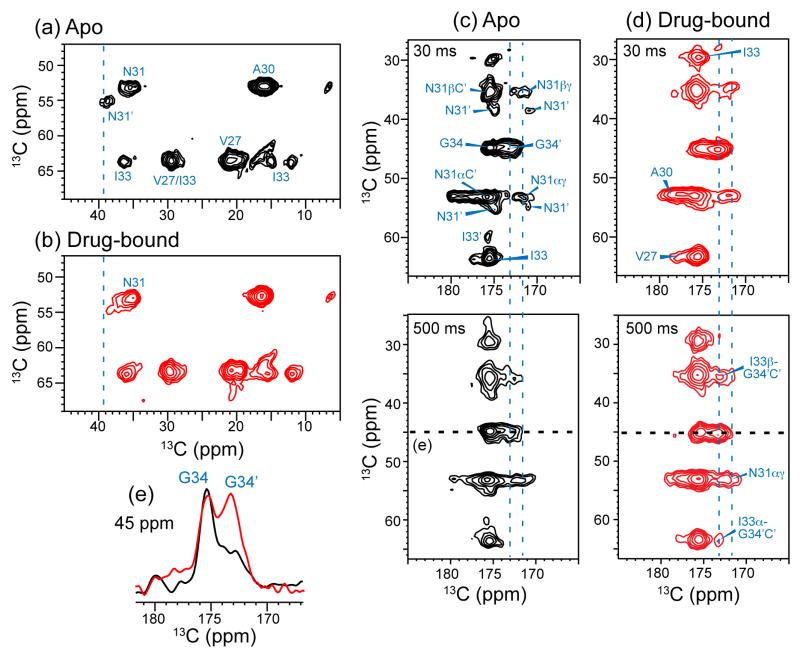
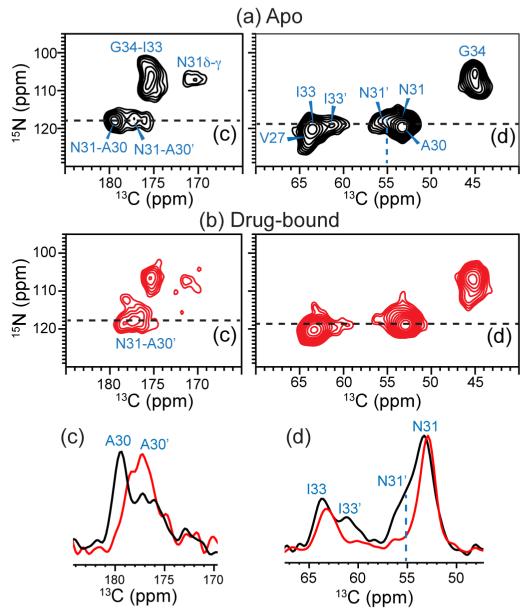
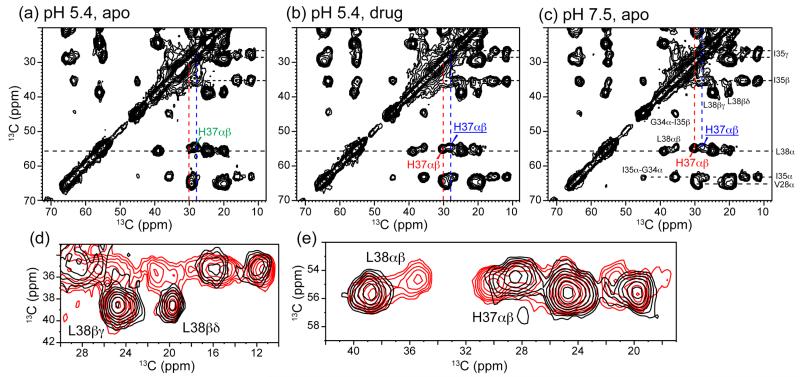
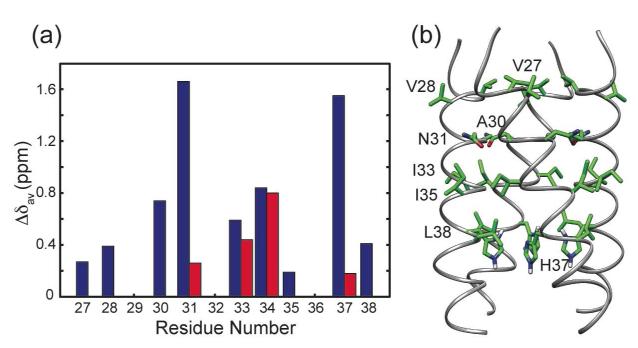
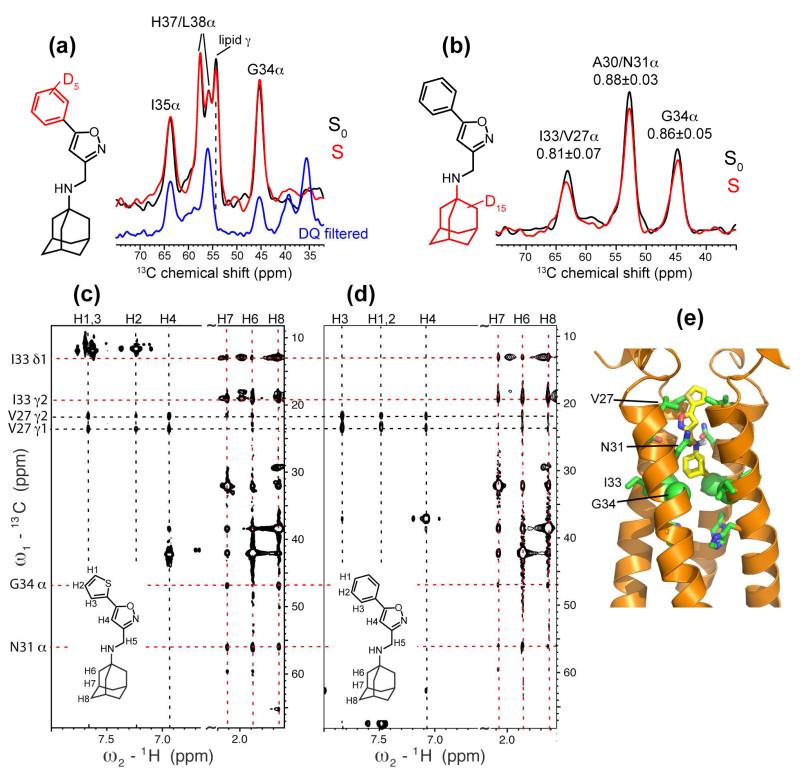
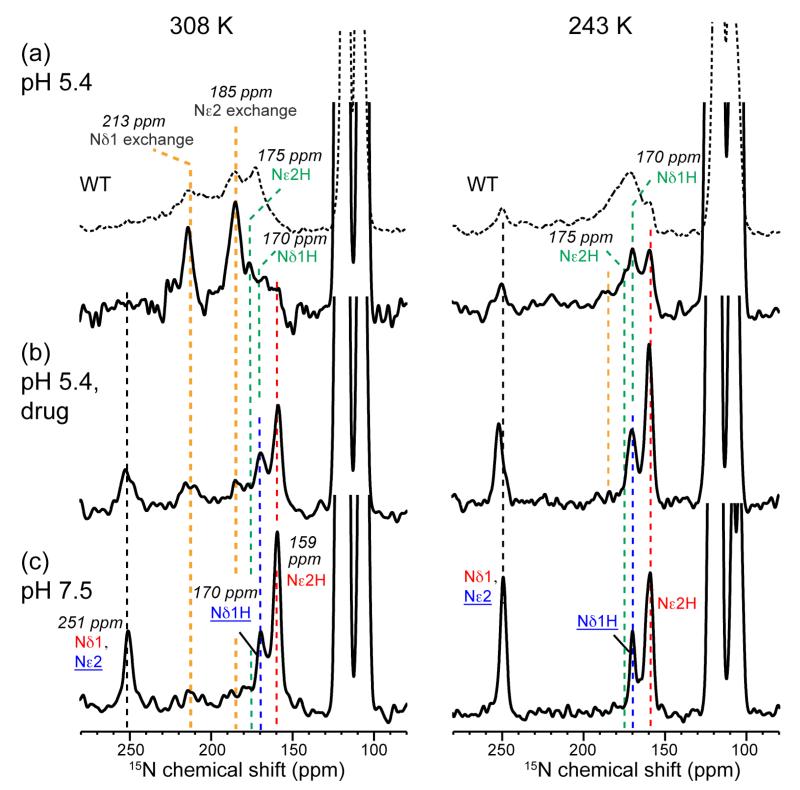
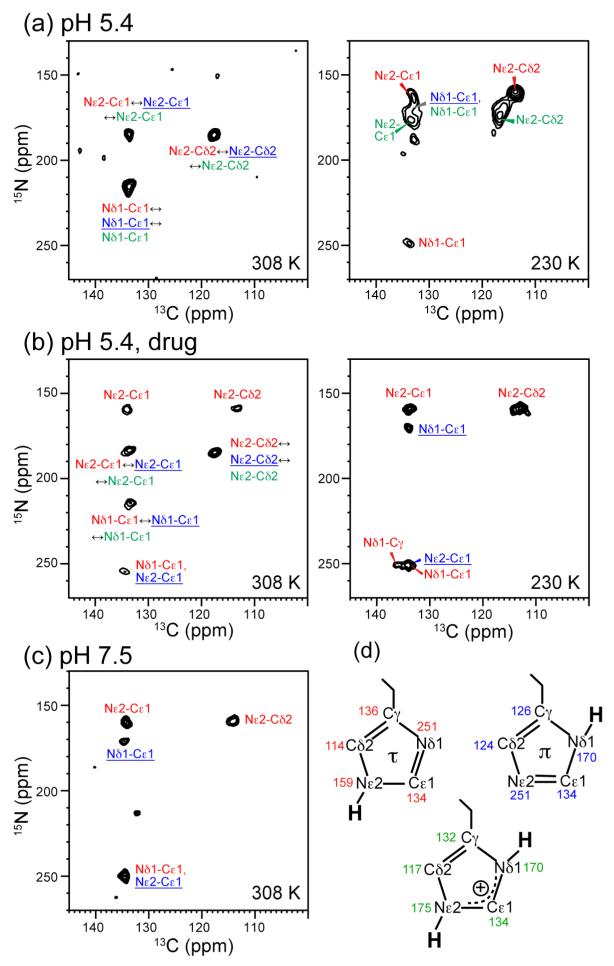
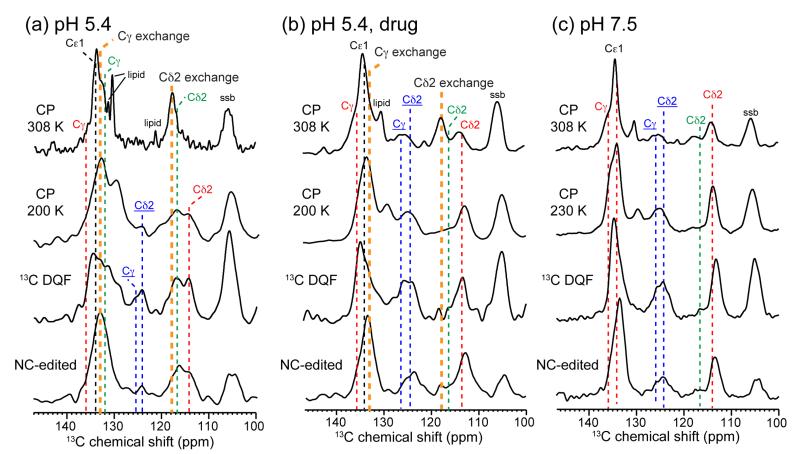
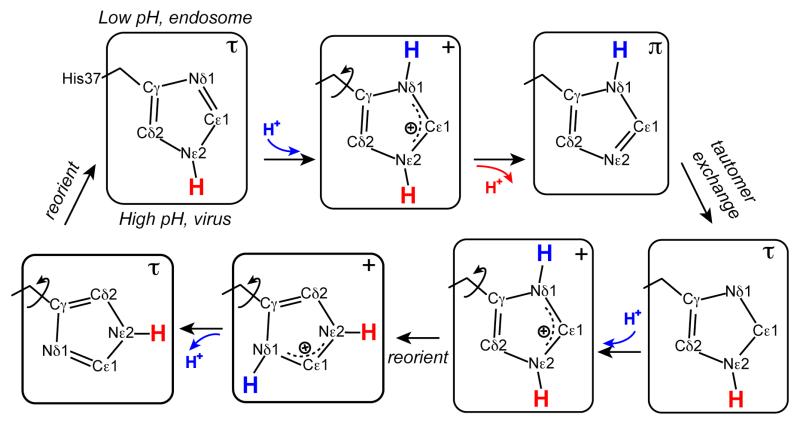
The M2 protein of influenza A viruses forms a tetrameric proton channel that is targeted by the amantadine class of antiviral drugs. A S31N mutation in the transmembrane (TM) domain of the protein has caused widespread amantadine resistance in most of the currently circulating flu viruses. Recently, a new family of compounds based on amantadine- and aryl-substituted isoxazole were discovered to inhibit the S31N channel activity and reduce replication of S31N-harboring viruses. We now use solid-state NMR spectroscopy to investigate the effects of one of these isoxazole compounds, WJ352, on the conformation of the S31N TM segment and the dynamics of the proton-selective residue, His37. Chemical shift perturbations show that WJ352 changes the conformational equilibrium of multiple TM residues, with the maximal perturbation occurring at the crucial Asn31. (13)C-(2)H distance measurements and (1)H-(1)H NOE cross peaks indicate that the adamantane moiety of the drug is bound in the spacious pore between Asn31 and Gly34 while the phenyl tail is located near Val27. Thus, the polar amine points to the channel exterior rather than to His37, in contrast to amantadine and rimantadine in the wild-type channel, suggesting that the drug is significantly stabilized by hydrophobic interactions between the adamantane and the TM peptide. (15)N and (13)C chemical shifts indicate that at low pH, His37 undergoes fast exchange among the τ tautomer, the π tautomer, and the cationic state due to proton transfer with water. The exchange rate is higher than the wild-type channel, consistent with the larger single-channel conductance of the mutant. Drug binding at acidic pH largely suppresses this exchange, reverting the histidines to a similar charge distribution as that of the high-pH closed state.
Figures
References
-
- Lamb RA, Holsinger KJ, Pinto LH. In: Cellular Receptors of Animal Viruses. Wemmer E, editor. Cold Spring Harbor Lab Press; Plainview, NY: 1994. pp. 303–321.
-
- Pinto LH, Lamb RA. J. Biol. Chem. 2006;281:8997–9000. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
