

Eliminating malaria vectors
- PMID: 23758937
- PMCID: PMC3685528
- DOI: 10.1186/1756-3305-6-172
Eliminating malaria vectors
Abstract
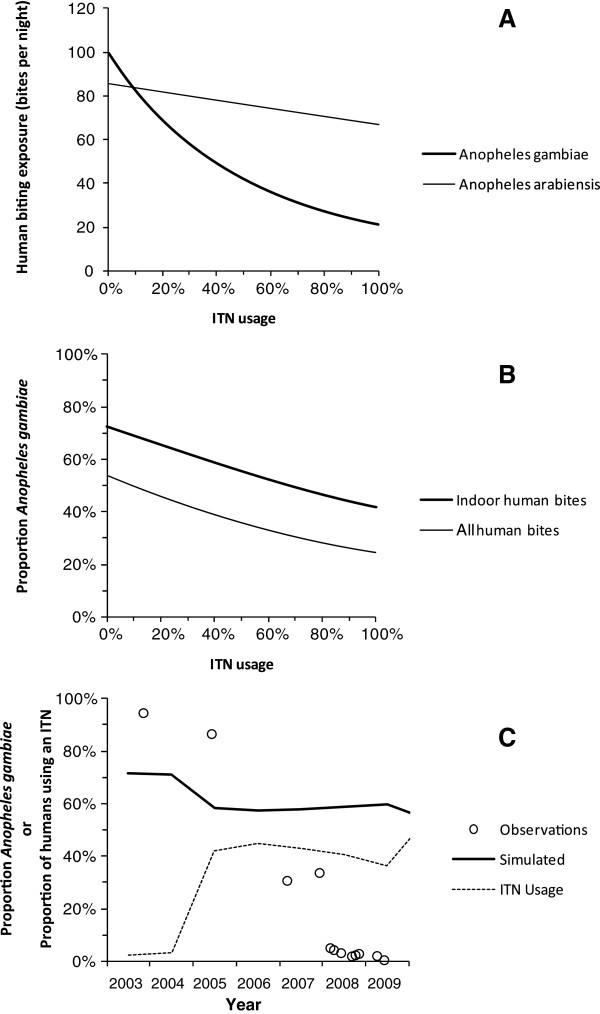
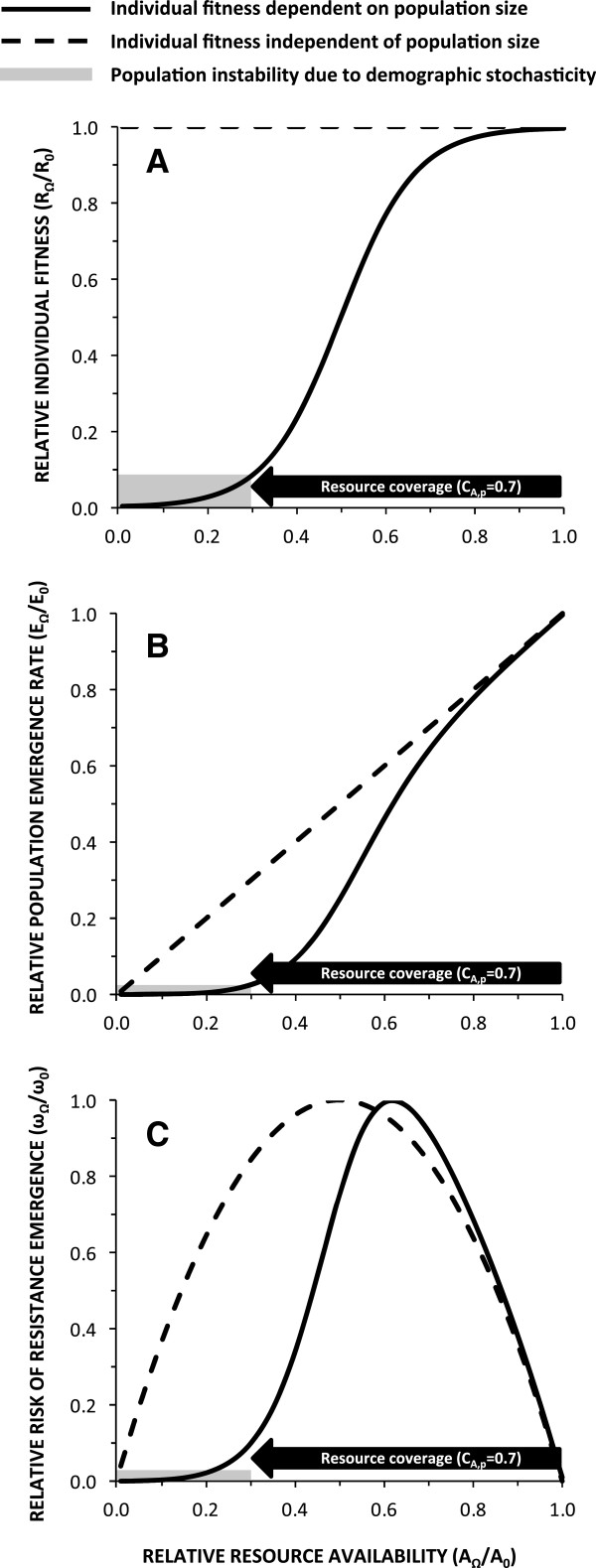
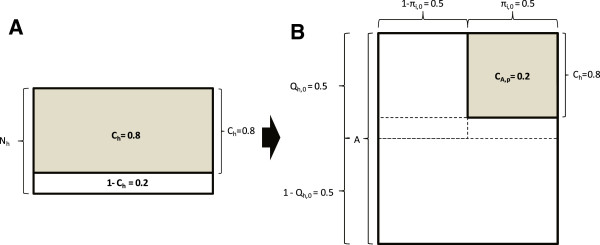
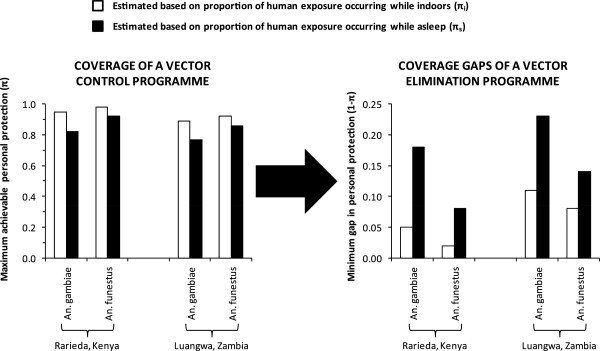
Malaria vectors which predominantly feed indoors upon humans have been locally eliminated from several settings with insecticide treated nets (ITNs), indoor residual spraying or larval source management. Recent dramatic declines of An. gambiae in east Africa with imperfect ITN coverage suggest mosquito populations can rapidly collapse when forced below realistically achievable, non-zero thresholds of density and supporting resource availability. Here we explain why insecticide-based mosquito elimination strategies are feasible, desirable and can be extended to a wider variety of species by expanding the vector control arsenal to cover a broader spectrum of the resources they need to survive. The greatest advantage of eliminating mosquitoes, rather than merely controlling them, is that this precludes local selection for behavioural or physiological resistance traits. The greatest challenges are therefore to achieve high biological coverage of targeted resources rapidly enough to prevent local emergence of resistance and to then continually exclude, monitor for and respond to re-invasion from external populations.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
