

Mitochondrial calcium uptake
- PMID: 23759742
- PMCID: PMC3696793
- DOI: 10.1073/pnas.1300410110
Mitochondrial calcium uptake
Abstract
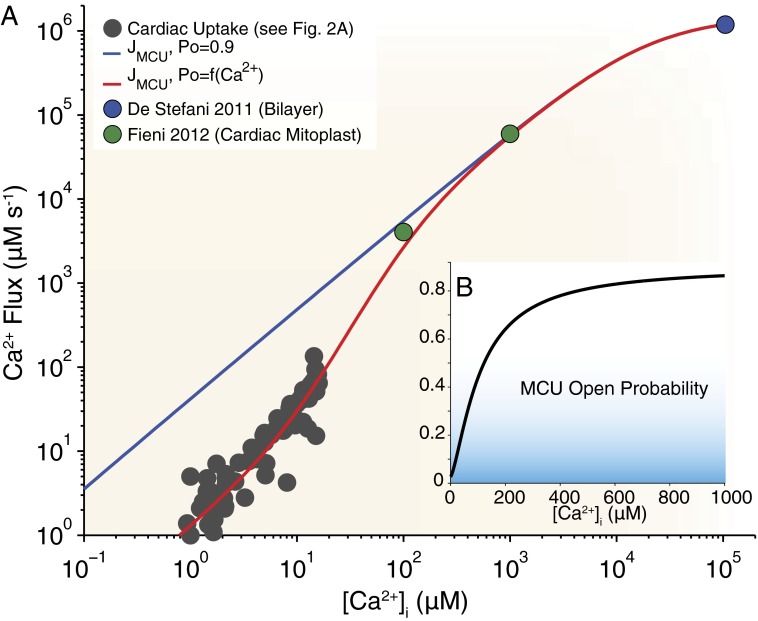
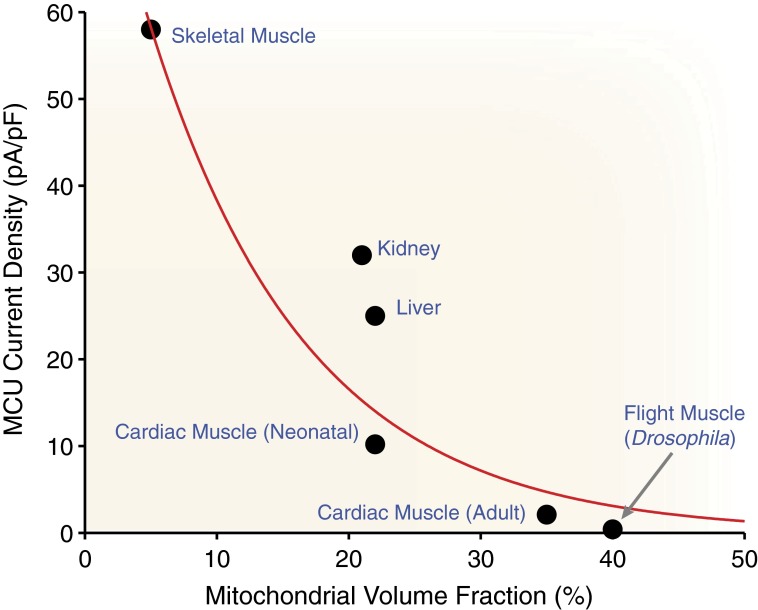
Calcium (Ca(2+)) uptake into the mitochondrial matrix is critically important to cellular function. As a regulator of matrix Ca(2+) levels, this flux influences energy production and can initiate cell death. If large, this flux could potentially alter intracellular Ca(2+) ([Ca(2+)]i) signals. Despite years of study, fundamental disagreements on the extent and speed of mitochondrial Ca(2+) uptake still exist. Here, we review and quantitatively analyze mitochondrial Ca(2+) uptake fluxes from different tissues and interpret the results with respect to the recently proposed mitochondrial Ca(2+) uniporter (MCU) candidate. This quantitative analysis yields four clear results: (i) under physiological conditions, Ca(2+) influx into the mitochondria via the MCU is small relative to other cytosolic Ca(2+) extrusion pathways; (ii) single MCU conductance is ∼6-7 pS (105 mM [Ca(2+)]), and MCU flux appears to be modulated by [Ca(2+)]i, suggesting Ca(2+) regulation of MCU open probability (P(O)); (iii) in the heart, two features are clear: the number of MCU channels per mitochondrion can be calculated, and MCU probability is low under normal conditions; and (iv) in skeletal muscle and liver cells, uptake per mitochondrion varies in magnitude but total uptake per cell still appears to be modest. Based on our analysis of available quantitative data, we conclude that although Ca(2+) critically regulates mitochondrial function, the mitochondria do not act as a significant dynamic buffer of cytosolic Ca(2+) under physiological conditions. Nevertheless, with prolonged (superphysiological) elevations of [Ca(2+)]i, mitochondrial Ca(2+) uptake can increase 10- to 1,000-fold and begin to shape [Ca(2+)]i dynamics.
Keywords: NCLX; NCX; SERCA; inner mitochondrial membrane; microdomain.
Conflict of interest statement
The authors declare no conflict of interest.
Figures


References
-
- Denton RM, McCormack JG. The role of calcium in the regulation of mitochondrial metabolism. Biochem Soc Trans. 1980;8(3):266–268. - PubMed
-
- Rizzuto R, Simpson AW, Brini M, Pozzan T. Rapid changes of mitochondrial Ca2+ revealed by specifically targeted recombinant aequorin. Nature. 1992;358(6384):325–327. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
