

The evolution and development of vertebrate lateral line electroreceptors
- PMID: 23761476
- PMCID: PMC4988487
- DOI: 10.1242/jeb.082362
The evolution and development of vertebrate lateral line electroreceptors
Abstract
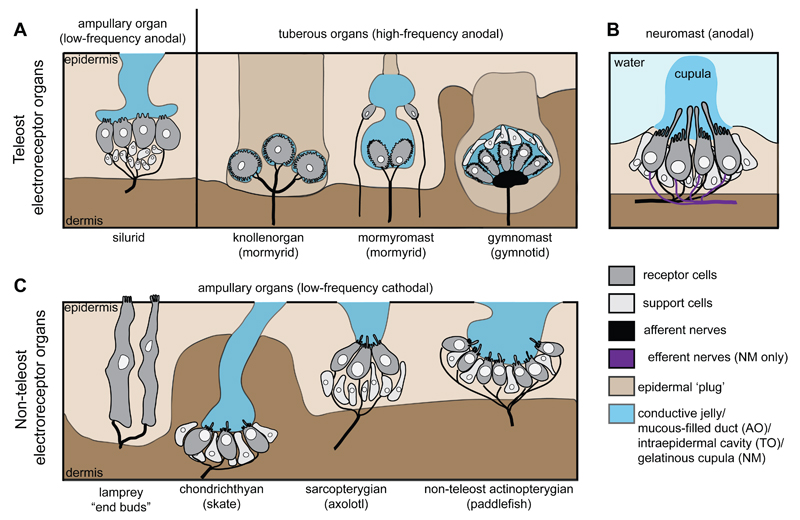
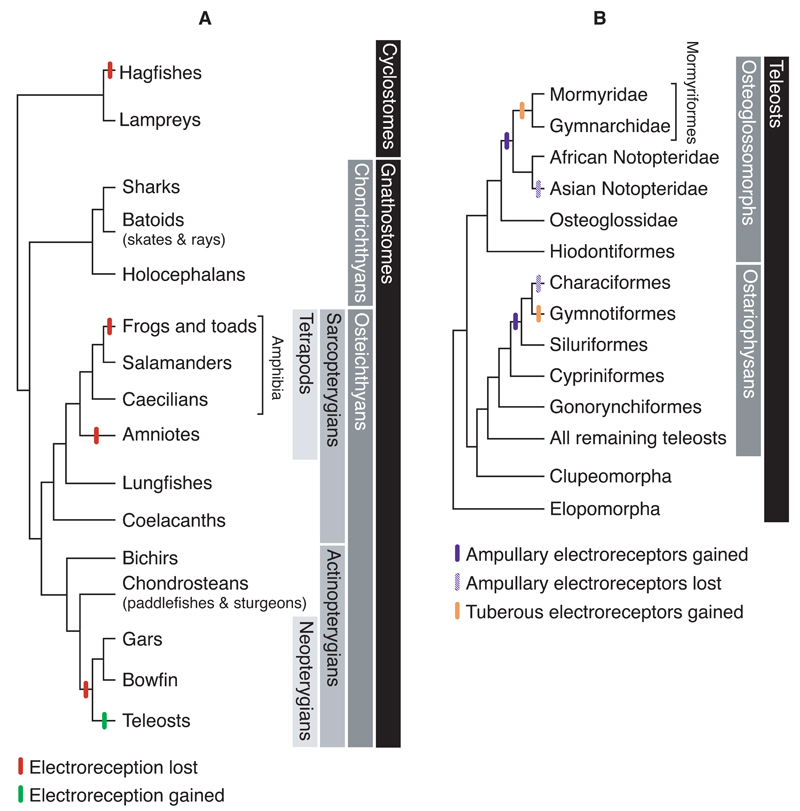
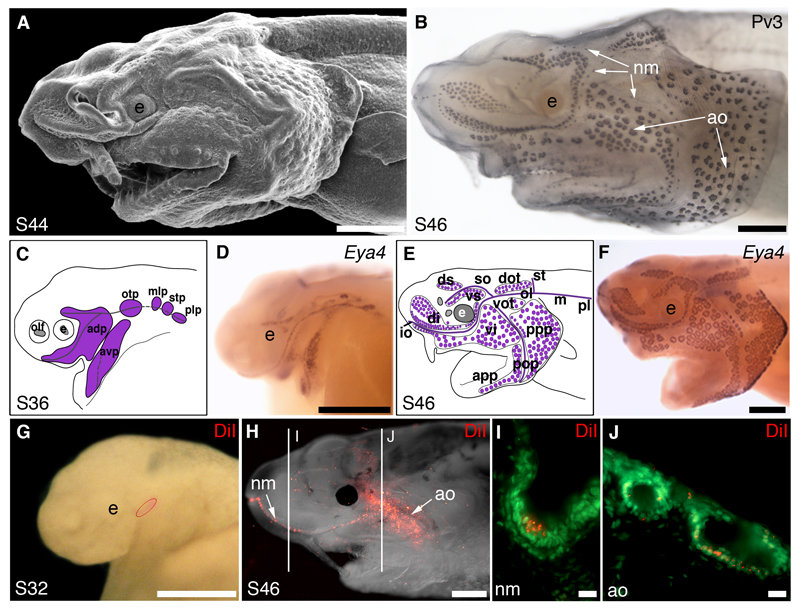
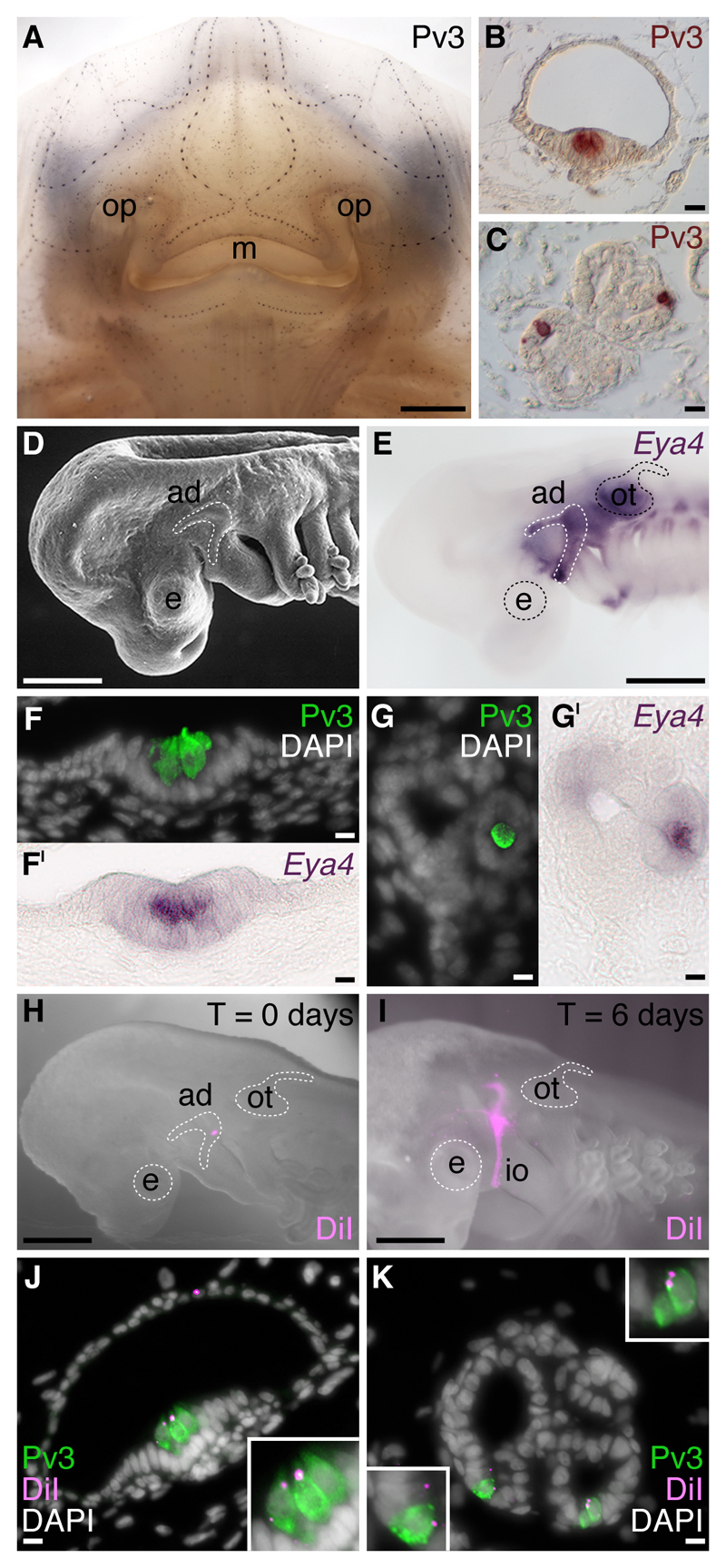
Electroreception is an ancient vertebrate sense with a fascinating evolutionary history involving multiple losses as well as independent evolution at least twice within teleosts. We review the phylogenetic distribution of electroreception and the morphology and innervation of electroreceptors in different vertebrate groups. We summarise recent work from our laboratory that has confirmed the homology of ampullary electroreceptors in non-teleost jawed vertebrates by showing, in conjunction with previously published work, that these are derived embryonically from lateral line placodes. Finally, we review hypotheses to explain the distribution of electroreception within teleosts, including the hypothesis that teleost ampullary and tuberous electroreceptors evolved via the modification of mechanosensory hair cells in lateral line neuromasts. We conclude that further experimental work on teleost electroreceptor development is needed to test such hypotheses.
Keywords: ampullary; electroreception; electroreceptors; hair cell; lateral line; neuromast; tuberous.
Figures
References
-
- Alves-Gomes JA. The evolution of electroreception and bioelectrogenesis in teleost fish: a phylogenetic perspective. J Fish Biol. 2001;58:1489–1511.
-
- Barry MA, White RL, Bennett MV. The elasmobranch spiracular organ. II. Physiological studies. J Comp Physiol A. 1988;163:93–98. - PubMed
-
- Baumann M, Roth A. The Ca++ permeability of the apical membrane of neuromast hair cells. J Comp Physiol A. 1986;158:681–688. - PubMed
-
- Bell CC, Maler L. Central neuroanatomy of electrosensory systems in fish. In: Bullock TH, Hopkins CD, Popper AN, Fay RR, editors. Electroreception. New York: Springer; 2005. pp. 68–111.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
