

Two-photon calcium imaging during fictive navigation in virtual environments
- PMID: 23761738
- PMCID: PMC3674334
- DOI: 10.3389/fncir.2013.00104
Two-photon calcium imaging during fictive navigation in virtual environments
Abstract
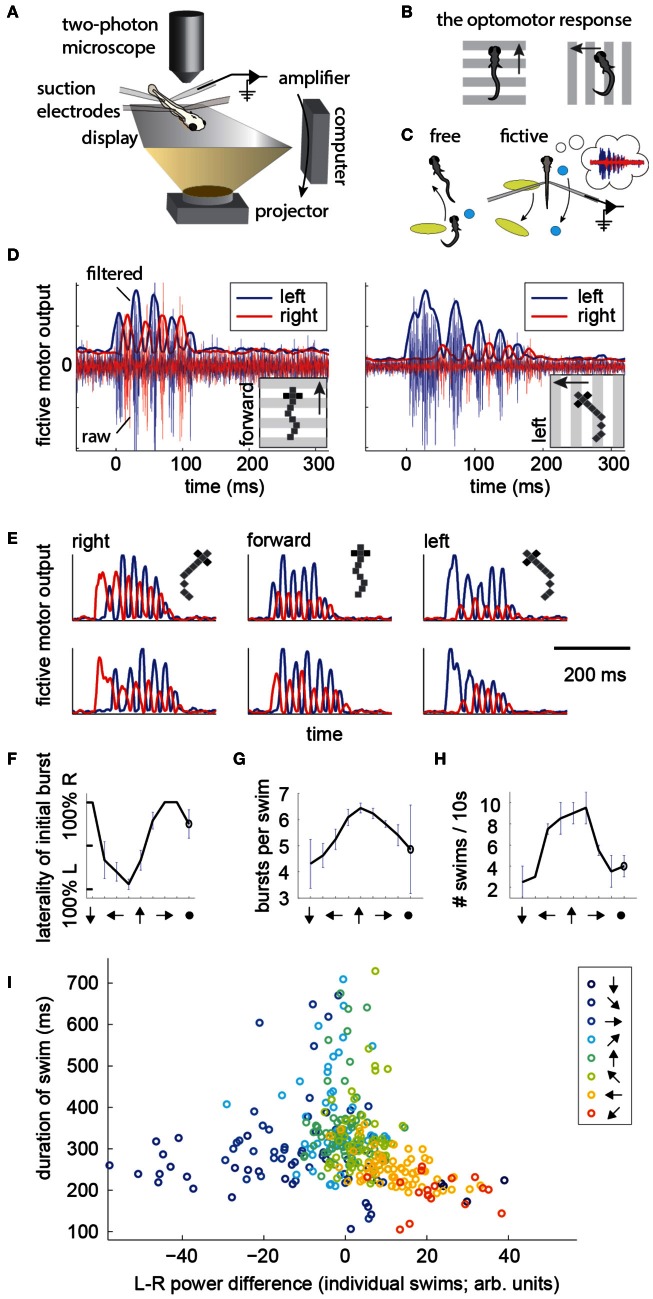
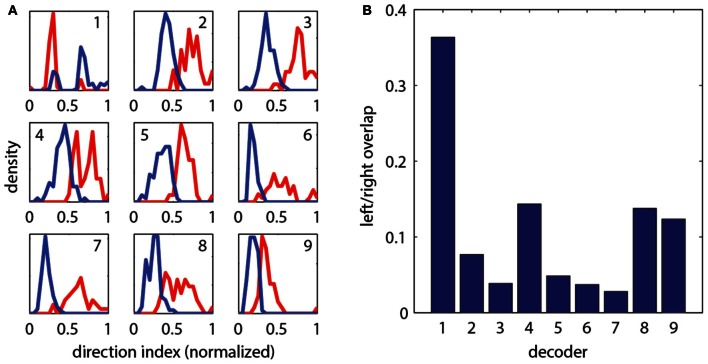
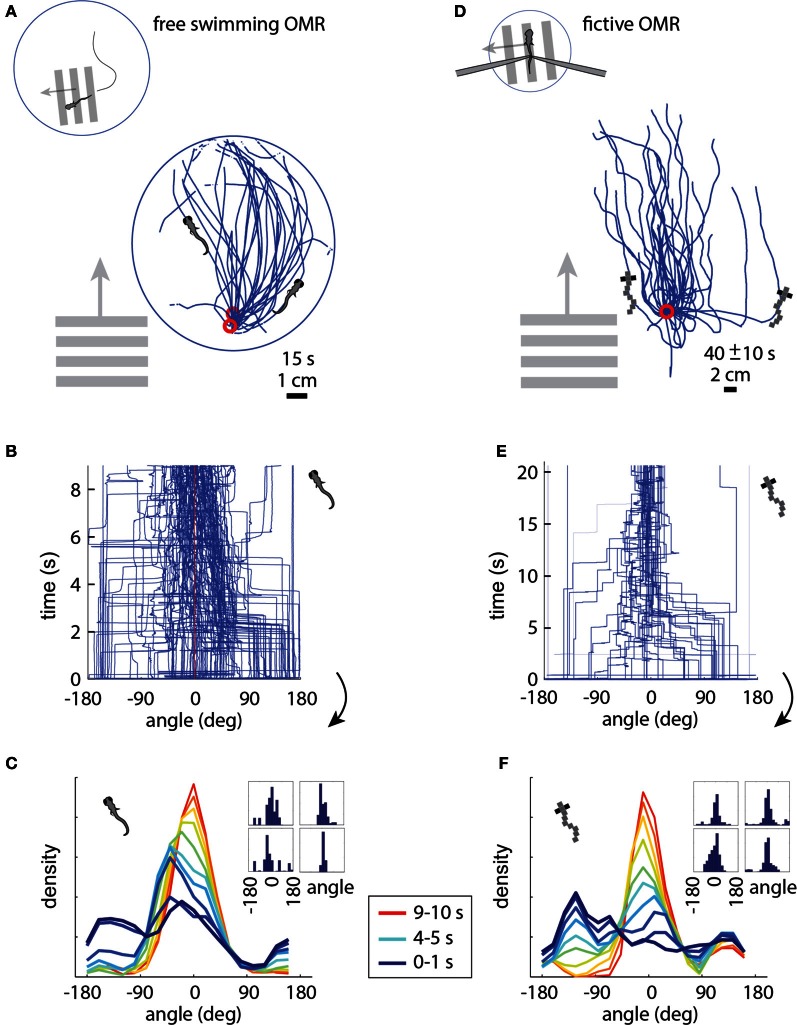
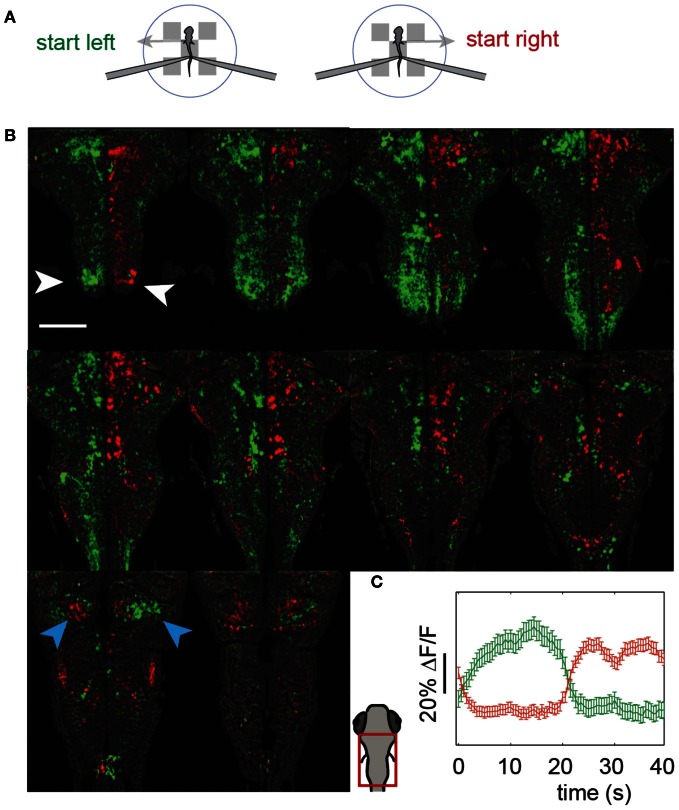
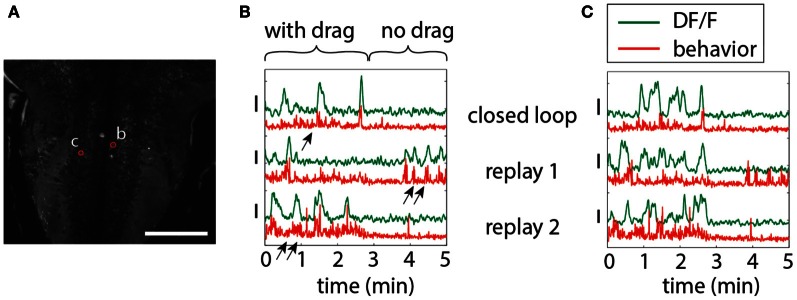
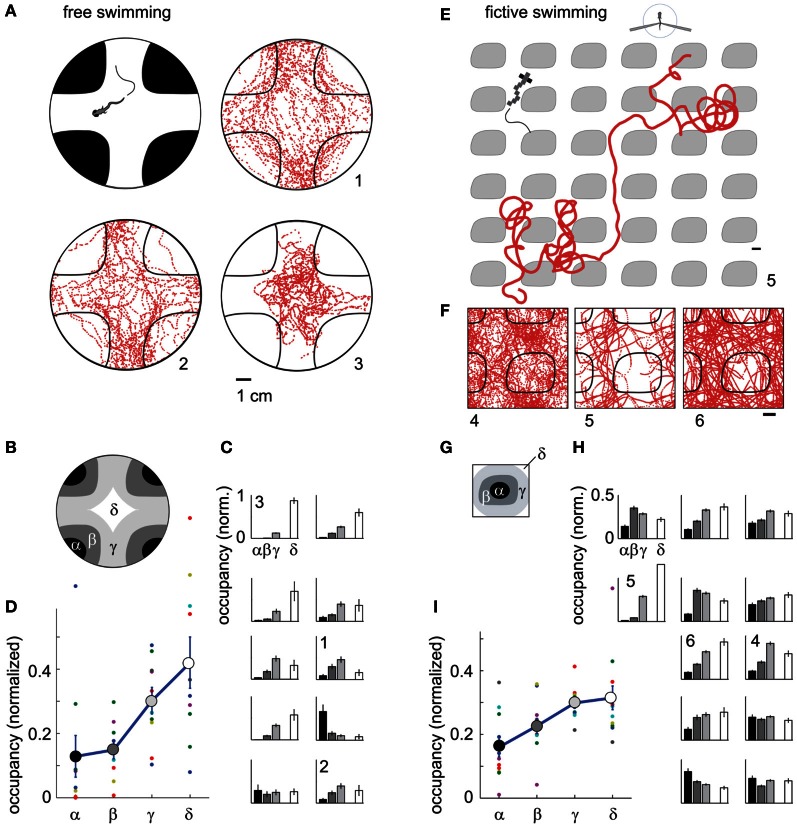
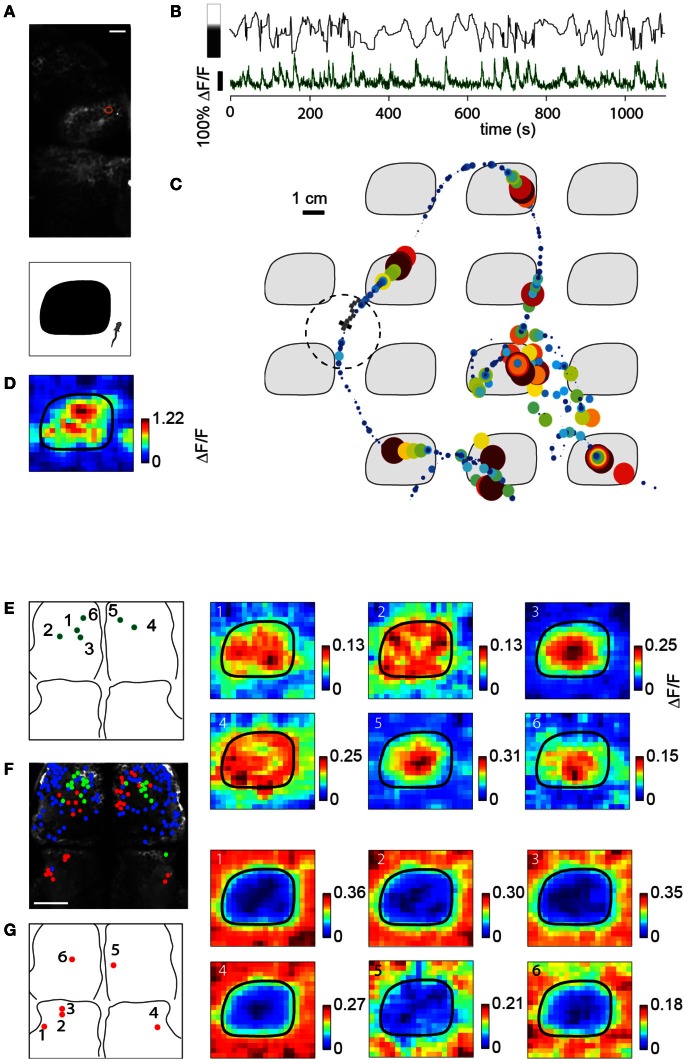
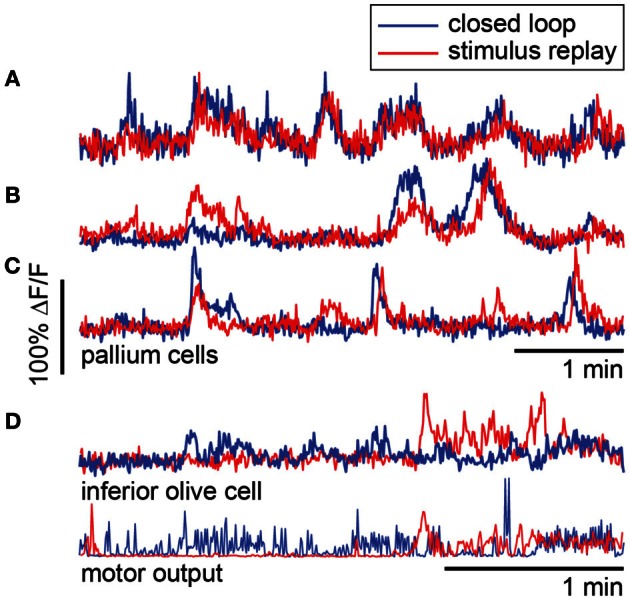
A full understanding of nervous system function requires recording from large populations of neurons during naturalistic behaviors. Here we enable paralyzed larval zebrafish to fictively navigate two-dimensional virtual environments while we record optically from many neurons with two-photon imaging. Electrical recordings from motor nerves in the tail are decoded into intended forward swims and turns, which are used to update a virtual environment displayed underneath the fish. Several behavioral features-such as turning responses to whole-field motion and dark avoidance-are well-replicated in this virtual setting. We readily observed neuronal populations in the hindbrain with laterally selective responses that correlated with right or left optomotor behavior. We also observed neurons in the habenula, pallium, and midbrain with response properties specific to environmental features. Beyond single-cell correlations, the classification of network activity in such virtual settings promises to reveal principles of brainwide neural dynamics during behavior.
Keywords: behavior; motor control; sensorimotor transformations; two-photon calcium imaging; virtual reality; zebrafish.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases