

Regulation of assimilate import into sink organs: update on molecular drivers of sink strength
- PMID: 23761804
- PMCID: PMC3671192
- DOI: 10.3389/fpls.2013.00177
Regulation of assimilate import into sink organs: update on molecular drivers of sink strength
Abstract
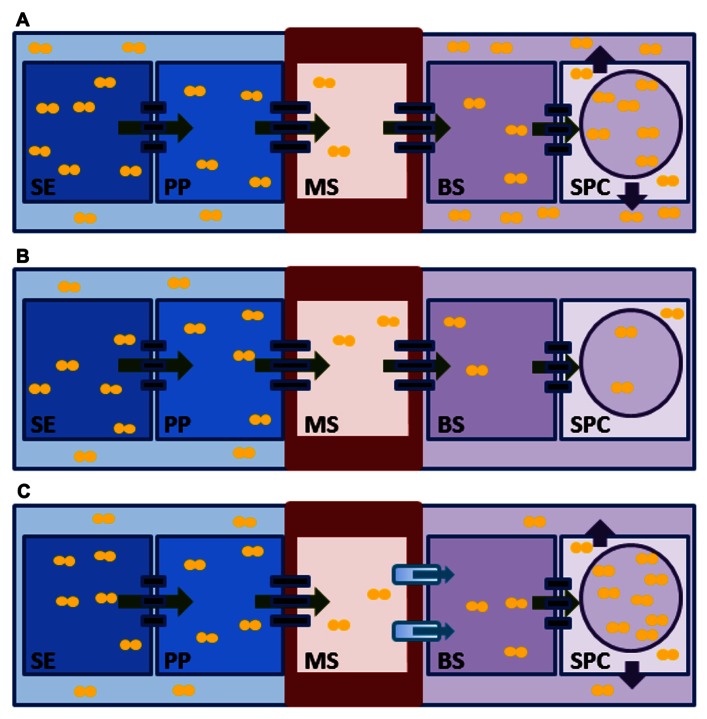
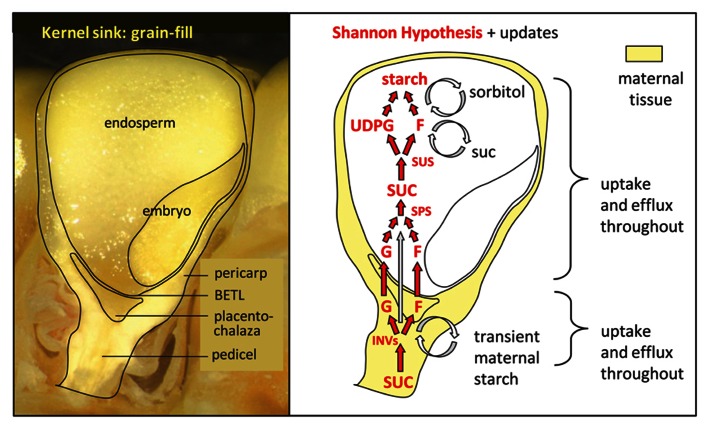
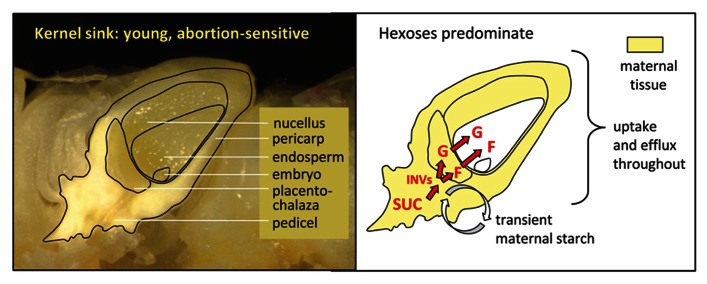
Recent developments have altered our view of molecular mechanisms that determine sink strength, defined here as the capacity of non-photosynthetic structures to compete for import of photoassimilates. We review new findings from diverse systems, including stems, seeds, flowers, and fruits. An important advance has been the identification of new transporters and facilitators with major roles in the accumulation and equilibration of sugars at a cellular level. Exactly where each exerts its effect varies among systems. Sugarcane and sweet sorghum stems, for example, both accumulate high levels of sucrose, but may do so via different paths. The distinction is central to strategies for targeted manipulation of sink strength using transporter genes, and shows the importance of system-specific analyses. Another major advance has been the identification of deep hypoxia as a feature of normal grain development. This means that molecular drivers of sink strength in endosperm operate in very low oxygen levels, and under metabolic conditions quite different than previously assumed. Successful enhancement of sink strength has nonetheless been achieved in grains by up-regulating genes for starch biosynthesis. Additionally, our understanding of sink strength is enhanced by awareness of the dual roles played by invertases (INVs), not only in sucrose metabolism, but also in production of the hexose sugar signals that regulate cell cycle and cell division programs. These contributions of INV to cell expansion and division prove to be vital for establishment of young sinks ranging from flowers to fruit. Since INV genes are themselves sugar-responsive "feast genes," they can mediate a feed-forward enhancement of sink strength when assimilates are abundant. Greater overall productivity and yield have thus been attained in key instances, indicating that even broader enhancements may be achievable as we discover the detailed molecular mechanisms that drive sink strength in diverse systems.
Keywords: carbohydrate partitioning; kernel; maize; sink strength; sorghum; stem; sucrose; sugarcane.
Figures
References
-
- Acreche M. M., Slafer G. A. (2006). Grain weight response to increases in number of grains in wheat in a Mediterranean area. Field Crops Res. 98 52–59 10.1016/j.fcr.2005.12.005 - DOI
-
- Ali M. L., Rajewski J. F., Baenziger P. S., Gill K. S., Eskridge K. M., Dweikat I. (2008). Assessment of genetic diversity and relationship among a collection of US sweet sorghum germplasm by SSR markers. Mol. Breed. 21 497–509 10.1007/s11032-007-9149-z - DOI
-
- Almodares A., Hadi M. R. (2009). Production of bioethanol from sweet sorghum: a review. Afr. J. Agric. Res. 4 772–780
LinkOut - more resources
Full Text Sources
Other Literature Sources