

Conditional deletion of Atoh1 reveals distinct critical periods for survival and function of hair cells in the organ of Corti
- PMID: 23761906
- PMCID: PMC3682389
- DOI: 10.1523/JNEUROSCI.5606-12.2013
Conditional deletion of Atoh1 reveals distinct critical periods for survival and function of hair cells in the organ of Corti
Abstract
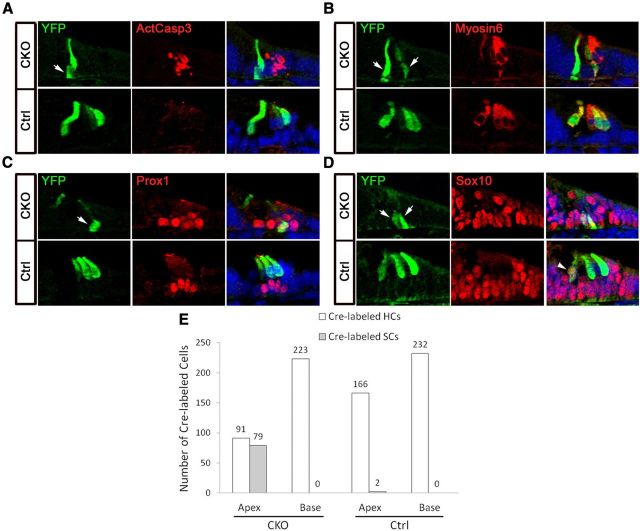
Atonal homolog1 (Atoh1) encodes a basic helix-loop-helix protein that is the first transcription factor to be expressed in differentiating hair cells. Previous work suggests that expression of Atoh1 in prosensory precursors is necessary for the differentiation and survival of hair cells, but it is not clear whether Atoh1 is required exclusively for these processes, or whether it regulates other functions later during hair cell maturation. We used EGFP-tagged Atoh1 knock-in mice to demonstrate for the first time that Atoh1 protein is expressed in hair cell precursors several days before the appearance of differentiated markers, but not in the broad pattern expected of a proneural gene. We conditionally deleted Atoh1 at different points in hair cell development and observe a rapid onset of hair cell defects, suggesting that the Atoh1 protein is unstable in differentiating hair cells and is necessary through an extended phase of their differentiation. Conditional deletion of Atoh1 reveals multiple functions in hair cell survival, maturation of stereociliary bundles, and auditory function. We show the presence of distinct critical periods for Atoh1 in each of these functions, suggesting that Atoh1 may be directly regulating many aspects of hair cell function. Finally, we show that the supporting cell death that accompanies loss of Atoh1 in hair cells is likely caused by the abortive trans-differentiation of supporting cells into hair cells. Together our data suggest that Atoh1 regulates multiple aspects of hair cell development and function.
Figures







References
-
- Chen P, Johnson JE, Zoghbi HY, Segil N. The role of Math1 in inner ear development: uncoupling the establishment of the sensory primordium from hair cell fate determination. Development. 2002;129:2495–2505. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases