

Streptolysin O and its co-toxin NAD-glycohydrolase protect group A Streptococcus from Xenophagic killing
- PMID: 23762025
- PMCID: PMC3675196
- DOI: 10.1371/journal.ppat.1003394
Streptolysin O and its co-toxin NAD-glycohydrolase protect group A Streptococcus from Xenophagic killing
Abstract
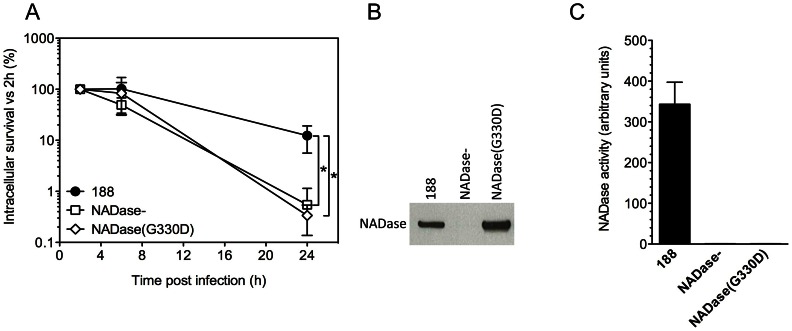
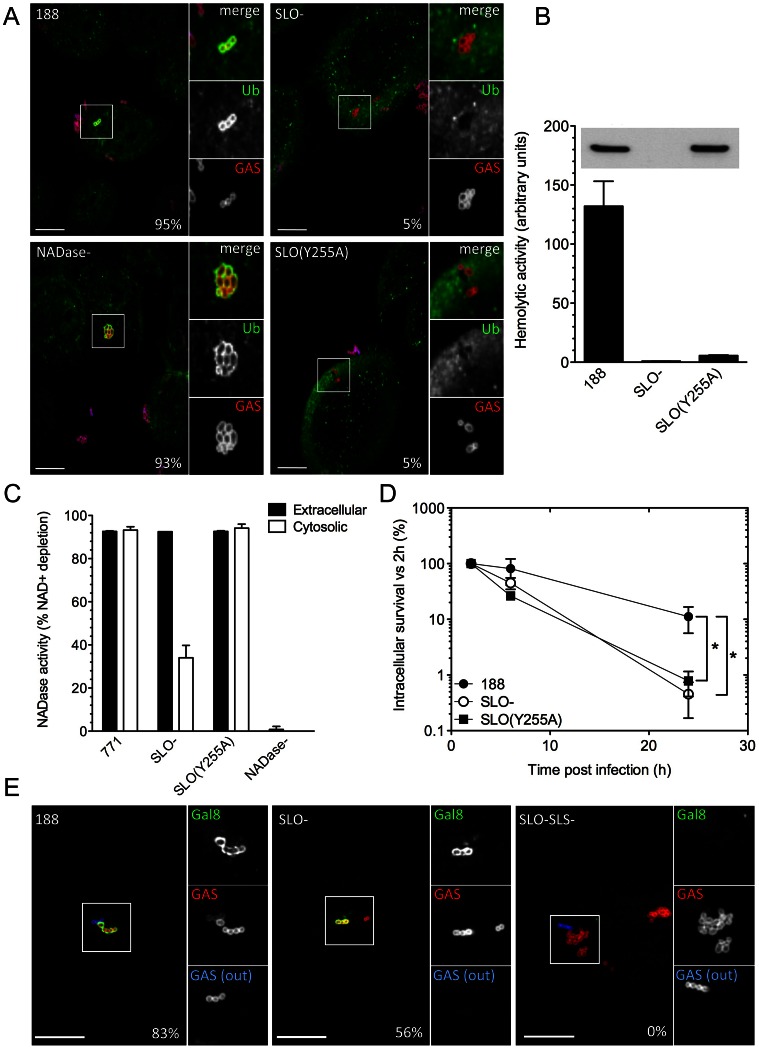
Group A Streptococcus (Streptococcus pyogenes or GAS) causes pharyngitis, severe invasive infections, and the post-infectious syndromes of glomerulonephritis and rheumatic fever. GAS can be internalized and killed by epithelial cells in vitro, a process that may contribute to local innate defense against pharyngeal infection. Secretion of the pore-forming toxin streptolysin O (SLO) by GAS has been reported to stimulate targeted autophagy (xenophagy) upon internalization of the bacteria by epithelial cells. Whereas this process was associated with killing of GAS in HeLa cells, studies in human keratinocytes found SLO production enhanced intracellular survival. To reconcile these conflicting observations, we now report in-depth investigation of xenophagy in response to GAS infection of human oropharyngeal keratinocytes, the predominant cell type of the pharyngeal epithelium. We found that SLO expression was associated with prolonged intracellular survival; unexpectedly, expression of the co-toxin NADase was required for this effect. Enhanced intracellular survival was lost upon deletion of NADase or inactivation of its enzymatic activity. Shortly after internalization of GAS by keratinocytes, SLO-mediated damage to the bacteria-containing vacuole resulted in exposure to the cytosol, ubiquitination of GAS and/or associated vacuolar membrane remnants, and engulfment of GAS in LC3-positive vacuoles. We also found that production of streptolysin S could mediate targeting of GAS to autophagosomes in the absence of SLO, a process accompanied by galectin 8 binding to damaged GAS-containing endosomes. Maturation of GAS-containing autophagosome-like vacuoles to degradative autolysosomes was prevented by SLO pore-formation and by SLO-mediated translocation of enzymatically active NADase into the keratinocyte cytosol. We conclude that SLO stimulates xenophagy in pharyngeal keratinocytes, but the coordinated action of SLO and NADase prevent maturation of GAS-containing autophagosomes, thereby prolonging GAS intracellular survival. This novel activity of NADase to block autophagic killing of GAS in pharyngeal cells may contribute to pharyngitis treatment failure, relapse, and chronic carriage.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures




References
-
- Stevens DL (2000) Group A beta-hemolytic streptococci: virulence factors, pathogenesis, and spectrum of clinical infections. In: Stevens DL, Kaplan EL, editors. Streptococcal Infections: Clinical Aspects, Microbiology, and Molecular Pathogenesis. New York: Oxford University Press. pp. 19–36.
-
- Gerber MA, Tanz RR, Kabat W, Bell GL, Siddiqui B, et al. (1999) Potential mechanisms for failure to eradicate group A streptococci from the pharynx. Pediatrics 104: 911–917. - PubMed
-
- Pichichero ME, Casey JR, Mayes T, Francis AB, Marsocci SM, et al. (2000) Penicillin failure in streptococcal tonsillopharyngitis: causes and remedies. Pediatr Infect Dis J 19: 917–923. - PubMed
-
- Greco R, De Martino L, Donnarumma G, Conte MP, Seganti L, et al. (1995) Invasion of cultured human cells by Streptococcus pyogenes . Res Microbiol 146: 551–560. - PubMed
-
- Jadoun J, Ozeri V, Burstein E, Skutelsky E, Hanski E, et al. (1998) Protein F1 is required for efficient entry of Streptococcus pyogenes into epithelial cells. J Infect Dis 178: 147–158. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
