

A Saccharomyces cerevisiae assay system to investigate ligand/AdipoR1 interactions that lead to cellular signaling
- PMID: 23762377
- PMCID: PMC3676391
- DOI: 10.1371/journal.pone.0065454
A Saccharomyces cerevisiae assay system to investigate ligand/AdipoR1 interactions that lead to cellular signaling
Abstract
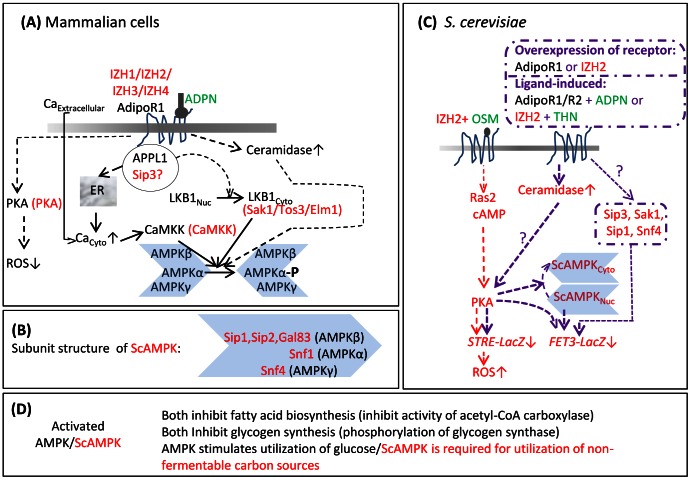
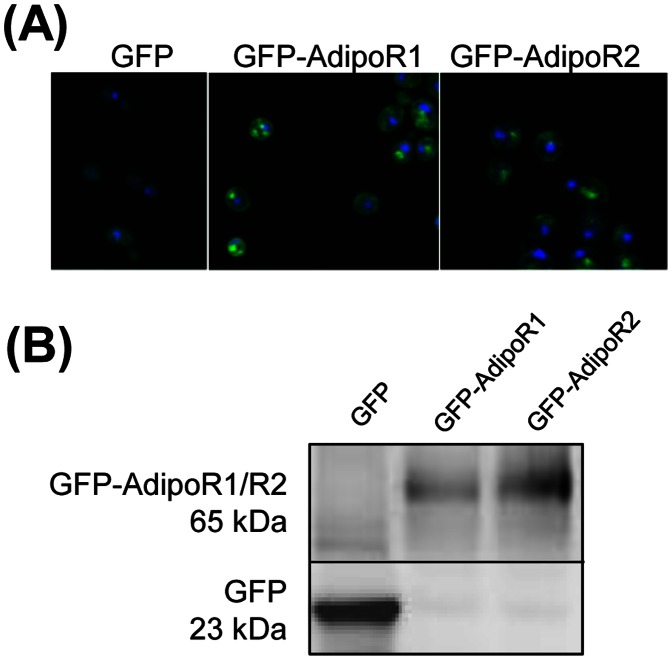
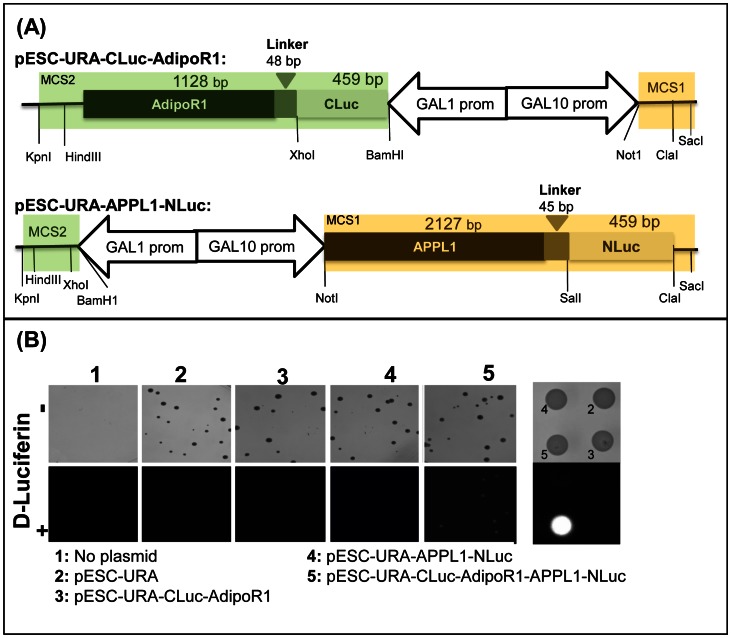
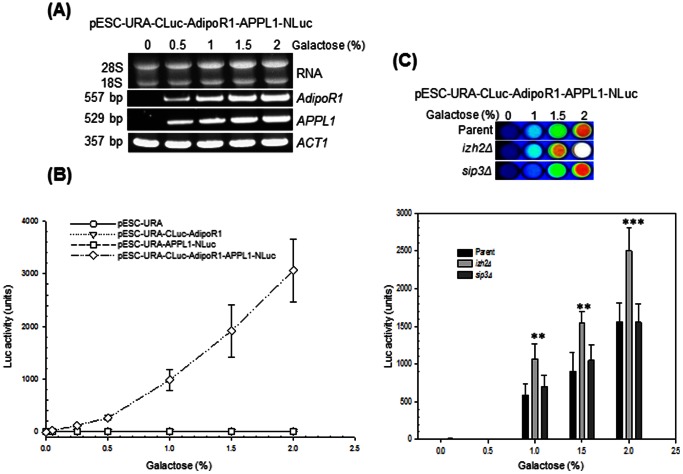
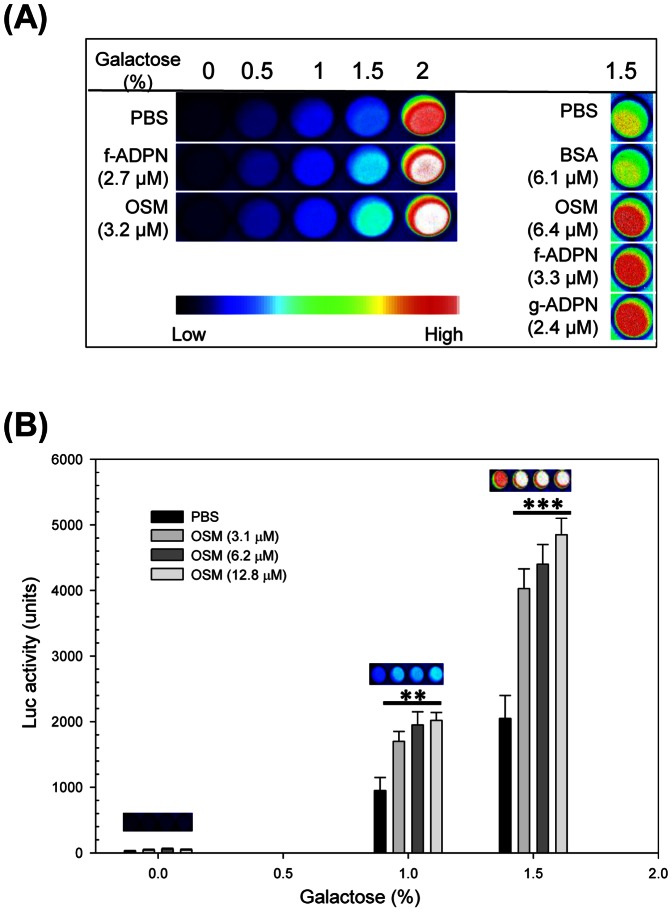
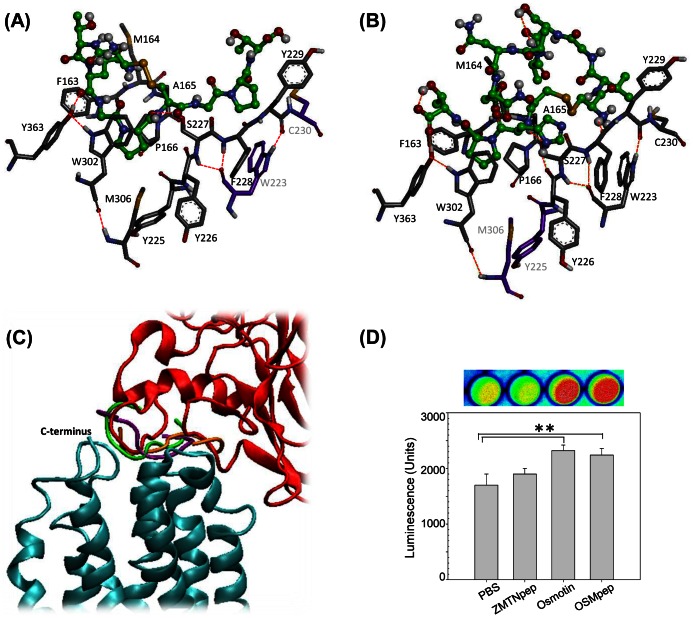
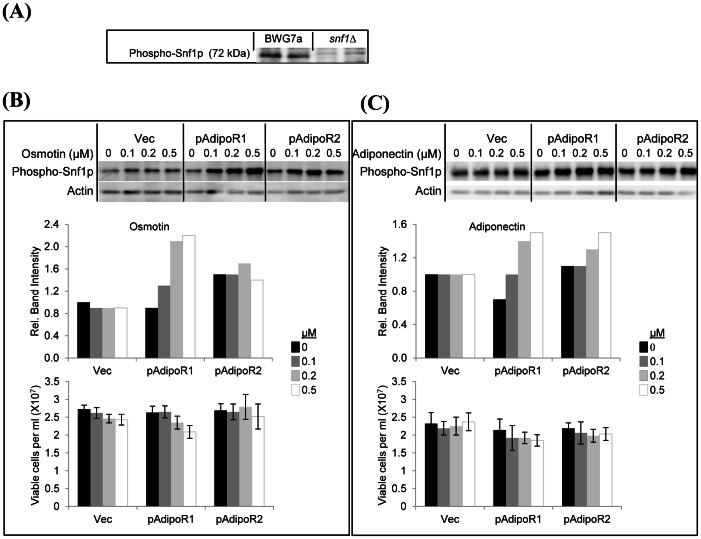
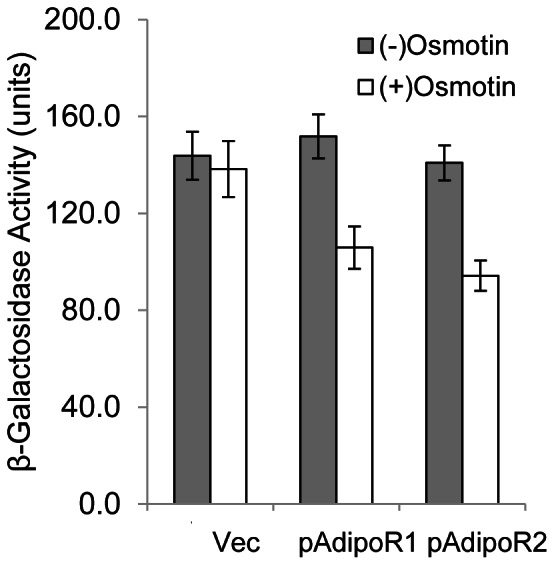
Adiponectin is a mammalian hormone that exerts anti-diabetic, anti-cancer and cardioprotective effects through interaction with its major ubiquitously expressed plasma membrane localized receptors, AdipoR1 and AdipoR2. Here, we report a Saccharomyces cerevisiae based method for investigating agonist-AdipoR interactions that is amenable for high-throughput scale-up and can be used to study both AdipoRs separately. Agonist-AdipoR1 interactions are detected using a split firefly luciferase assay based on reconstitution of firefly luciferase (Luc) activity due to juxtaposition of its N- and C-terminal fragments, NLuc and CLuc, by ligand induced interaction of the chimeric proteins CLuc-AdipoR1 and APPL1-NLuc (adaptor protein containing pleckstrin homology domain, phosphotyrosine binding domain and leucine zipper motif 1-NLuc) in a S. cerevisiae strain lacking the yeast homolog of AdipoRs (Izh2p). The assay monitors the earliest known step in the adiponectin-AdipoR anti-diabetic signaling cascade. We demonstrate that reconstituted Luc activity can be detected in colonies or cells using a CCD camera and quantified in cell suspensions using a microplate reader. AdipoR1-APPL1 interaction occurs in absence of ligand but can be stimulated specifically by agonists such as adiponectin and the tobacco protein osmotin that was shown to have AdipoR-dependent adiponectin-like biological activity in mammalian cells. To further validate this assay, we have modeled the three dimensional structures of receptor-ligand complexes of membrane-embedded AdipoR1 with cyclic peptides derived from osmotin or osmotin-like plant proteins. We demonstrate that the calculated AdipoR1-peptide binding energies correlate with the peptides' ability to behave as AdipoR1 agonists in the split luciferase assay. Further, we demonstrate agonist-AdipoR dependent activation of protein kinase A (PKA) signaling and AMP activated protein kinase (AMPK) phosphorylation in S. cerevisiae, which are homologous to important mammalian adiponectin-AdipoR1 signaling pathways. This system should facilitate the development of therapeutic inventions targeting adiponectin and/or AdipoR physiology.
Conflict of interest statement
Figures
References
-
- Berg AH, Combs TP, Scherer PE (2002) ACRP30/adiponectin: an adipokine regulating glucose and lipid metabolism. Trends Endocrinol Metab 13: 84–89. - PubMed
-
- Jortay J, Senou M, Abou-Samra M, Noel L, Robert A, et al. (2012) Adiponectin and skeletal muscle: pathophysiological implications in metabolic stress. Am J Pathol 181: 245–256. - PubMed
-
- Kadowaki T, Yamauchi T (2005) Adiponectin and adiponectin receptors. Endocr Rev 26: 439–451. - PubMed
-
- Hara K, Boutin P, Mori Y, Tobe K, Dina C, et al. (2002) Genetic variation in the gene encoding adiponectin is associated with an increased risk of type 2 diabetes in the Japanese population. Diabetes 51: 536–540. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
