

Impact of neuronal properties on network coding: roles of spike initiation dynamics and robust synchrony transfer
- PMID: 23764282
- PMCID: PMC3753823
- DOI: 10.1016/j.neuron.2013.05.030
Impact of neuronal properties on network coding: roles of spike initiation dynamics and robust synchrony transfer
Abstract
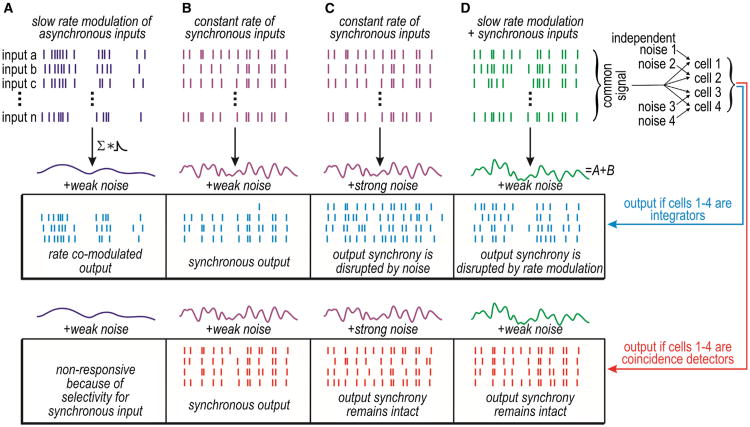
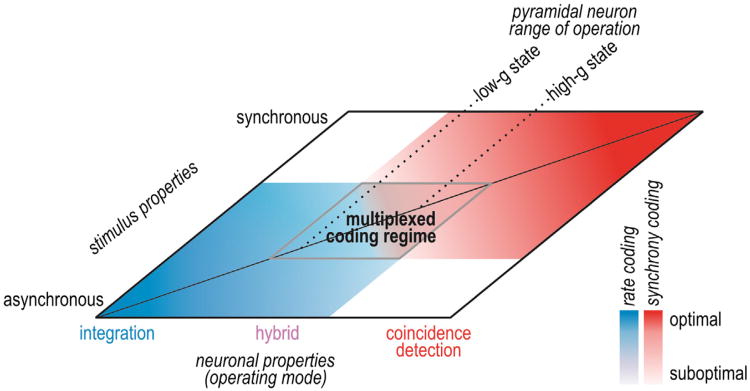
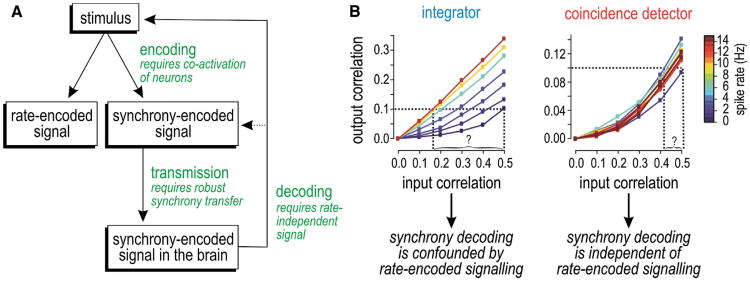
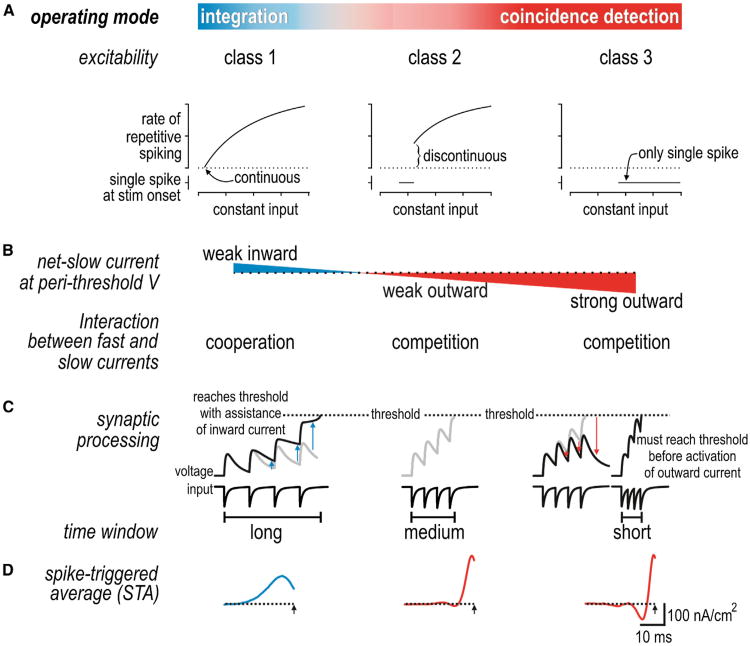
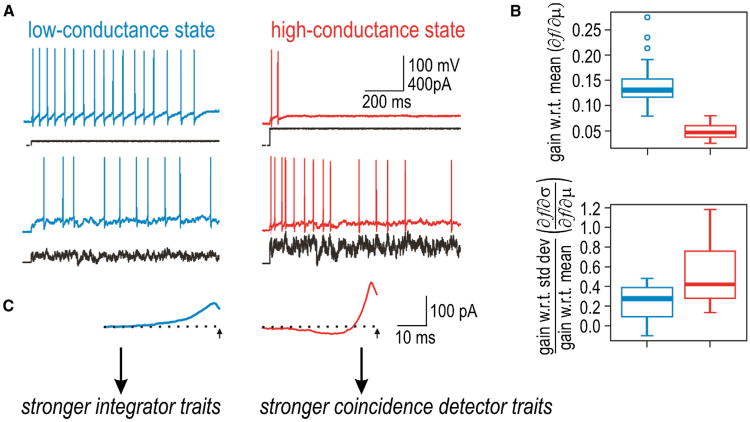
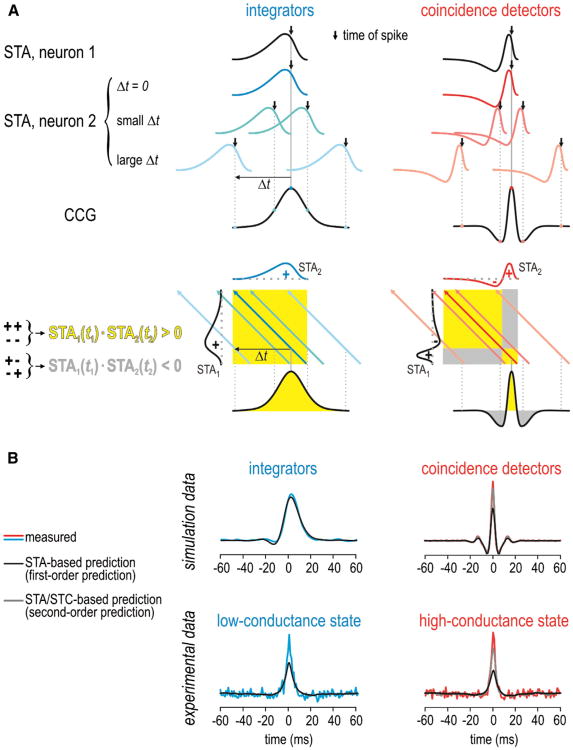
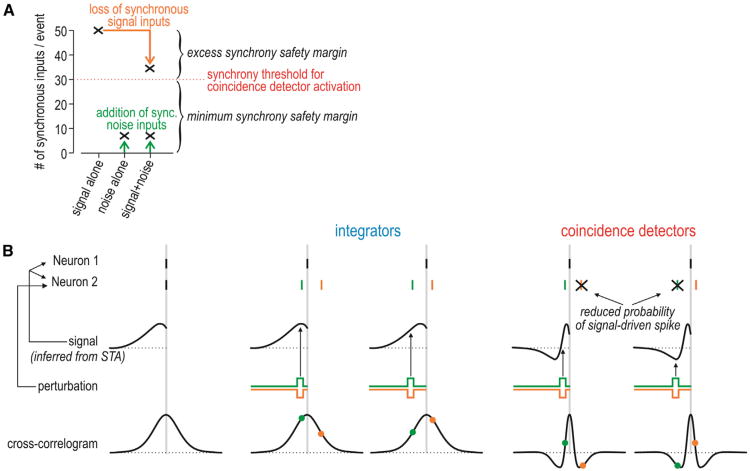
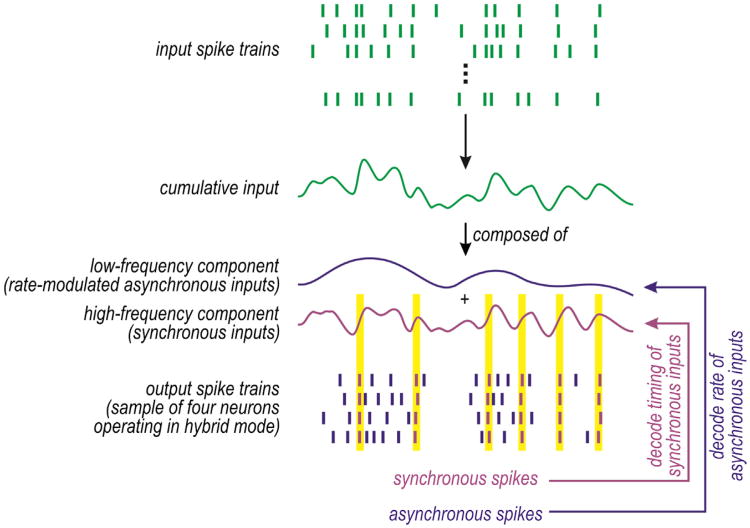
Neural networks are more than the sum of their parts, but the properties of those parts are nonetheless important. For instance, neuronal properties affect the degree to which neurons receiving common input will spike synchronously, and whether that synchrony will propagate through the network. Stimulus-evoked synchrony can help or hinder network coding depending on the type of code. In this Perspective, we describe how spike initiation dynamics influence neuronal input-output properties, how those properties affect synchronization, and how synchronization affects network coding. We propose that synchronous and asynchronous spiking can be used to multiplex temporal (synchrony) and rate coding and discuss how pyramidal neurons would be well suited for that task.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures
References
-
- Abeles M. Role of the cortical neuron: integrator or coincidence detector? Isr J Med Sci. 1982;18:83–92. - PubMed
-
- Agüera y Arcas B, Fairhall AL. What causes a neuron to spike? Neural Comput. 2003;15:1789–1807. - PubMed
-
- Agüera y Arcas B, Fairhall AL, Bialek W. Computation in a single neuron: Hodgkin and Huxley revisited. Neural Comput. 2003;15:1715–1749. - PubMed
-
- Alonso JM, Usrey WM, Reid RC. Precisely correlated firing in cells of the lateral geniculate nucleus. Nature. 1996;383:815–819. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
