

Differential expression of multidrug resistance protein 5 and phosphodiesterase 5 and regulation of cGMP levels in phasic and tonic smooth muscle
- PMID: 23764893
- PMCID: PMC3891211
- DOI: 10.1152/ajpgi.00457.2012
Differential expression of multidrug resistance protein 5 and phosphodiesterase 5 and regulation of cGMP levels in phasic and tonic smooth muscle
Abstract
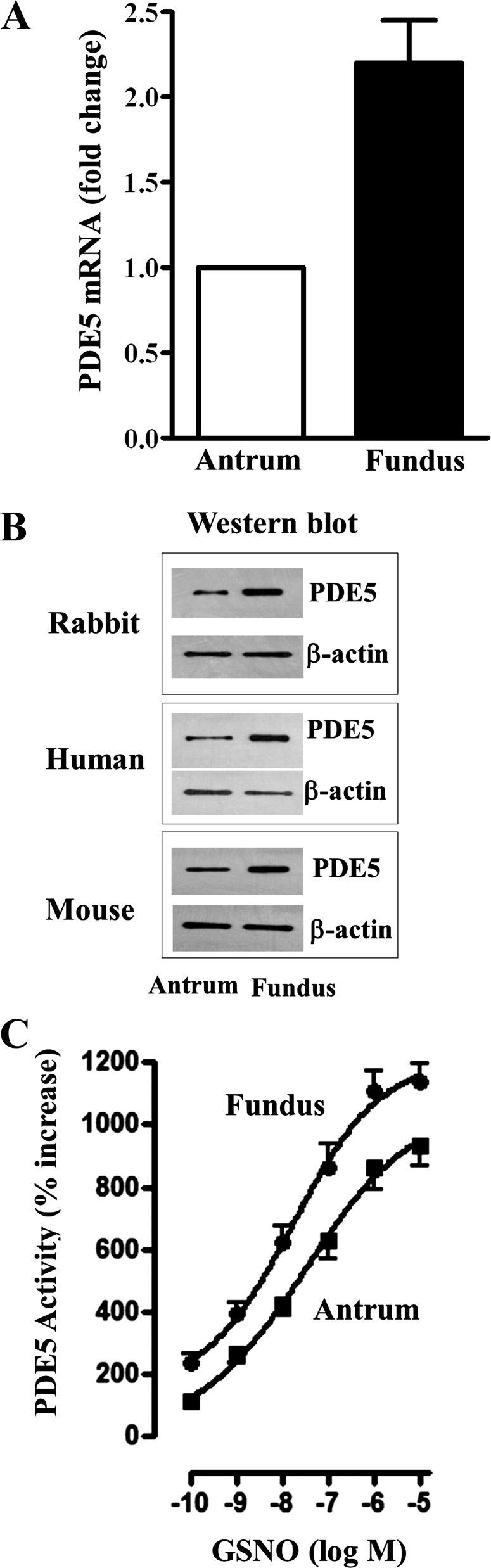
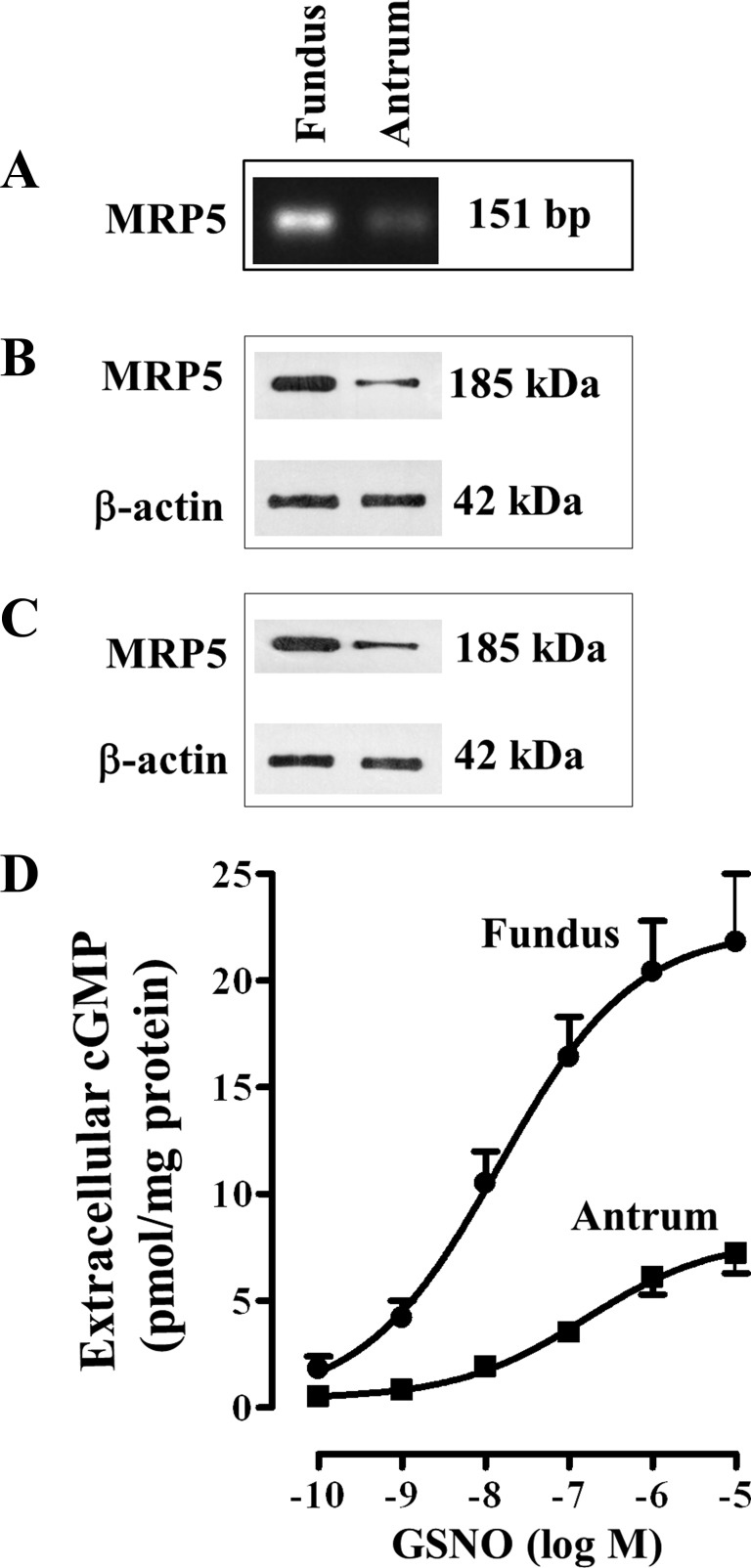
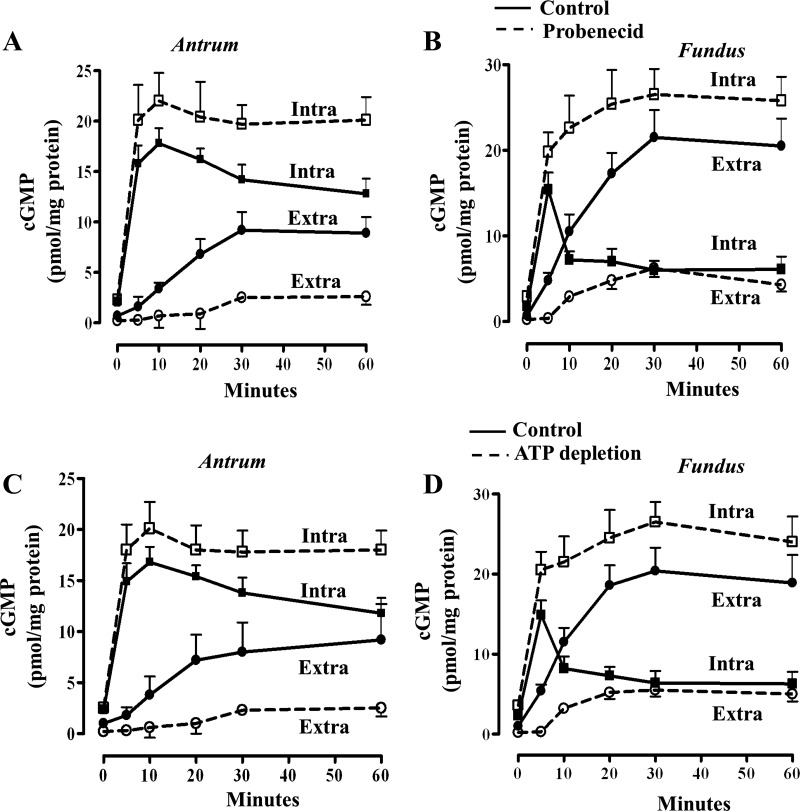
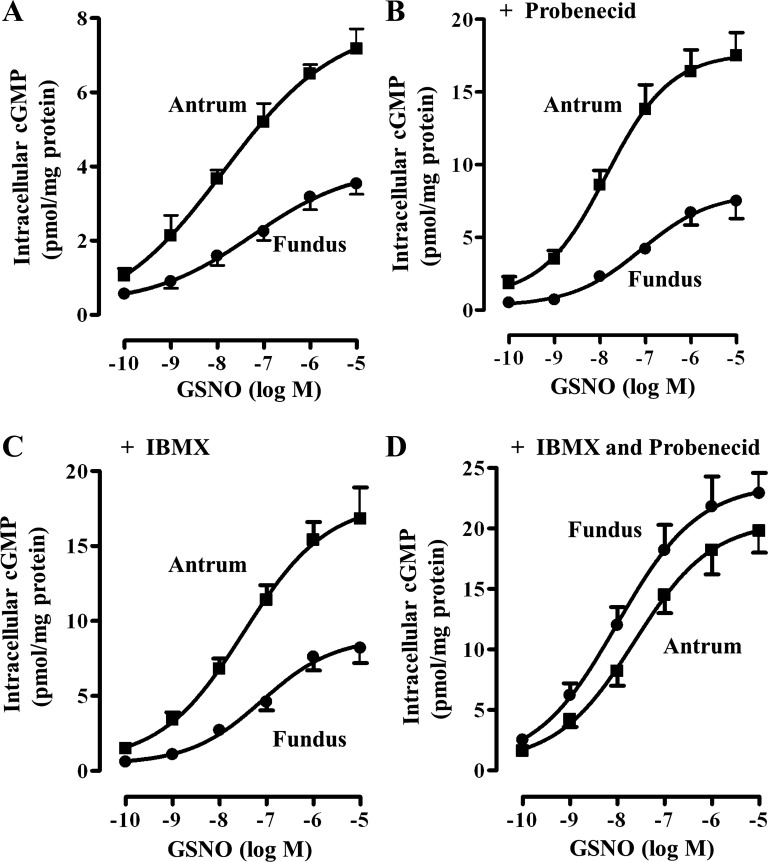
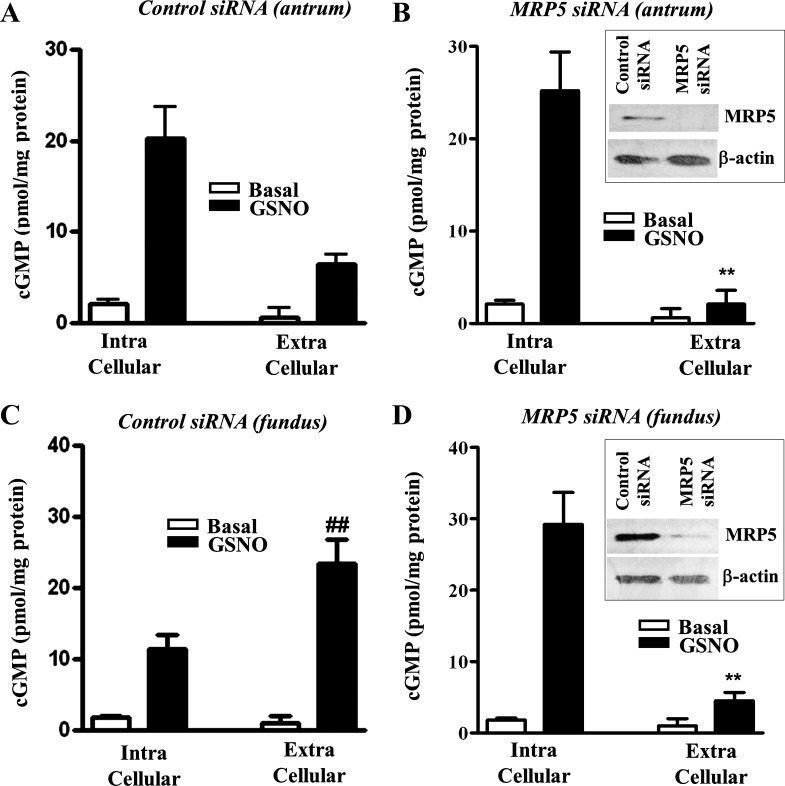
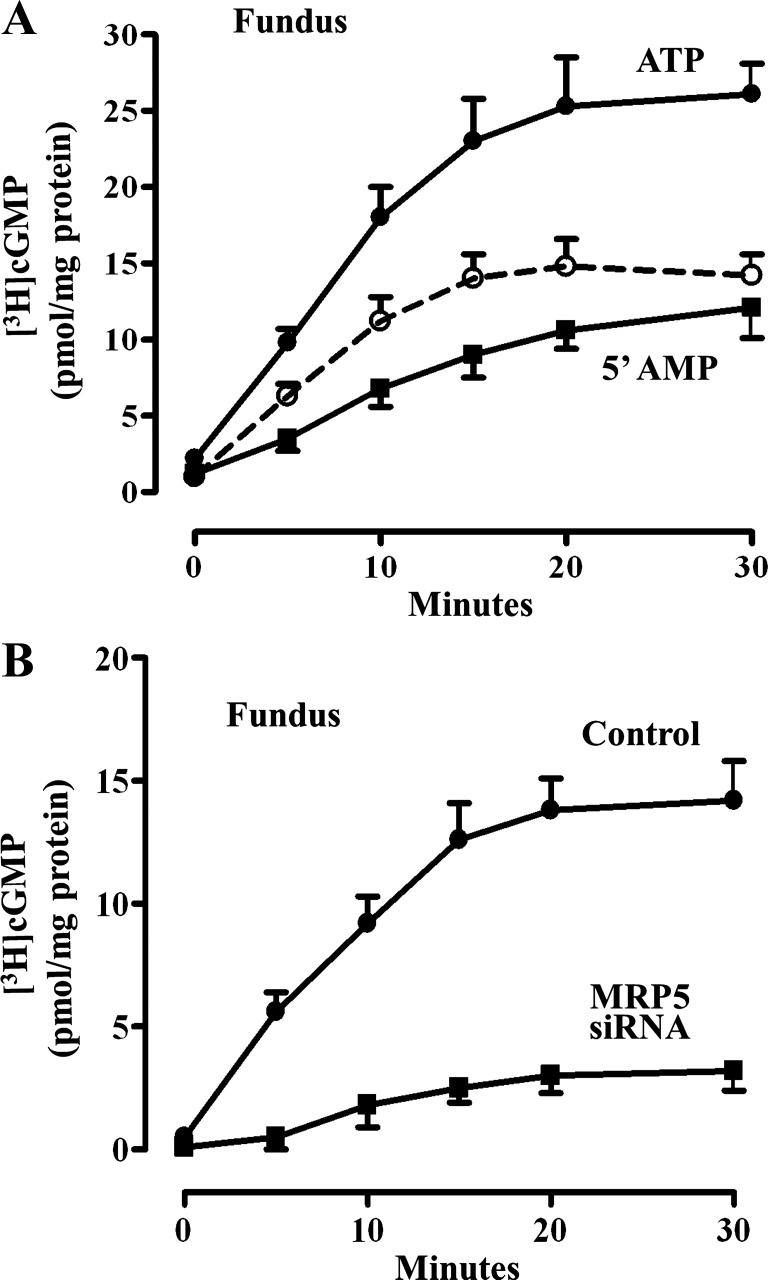
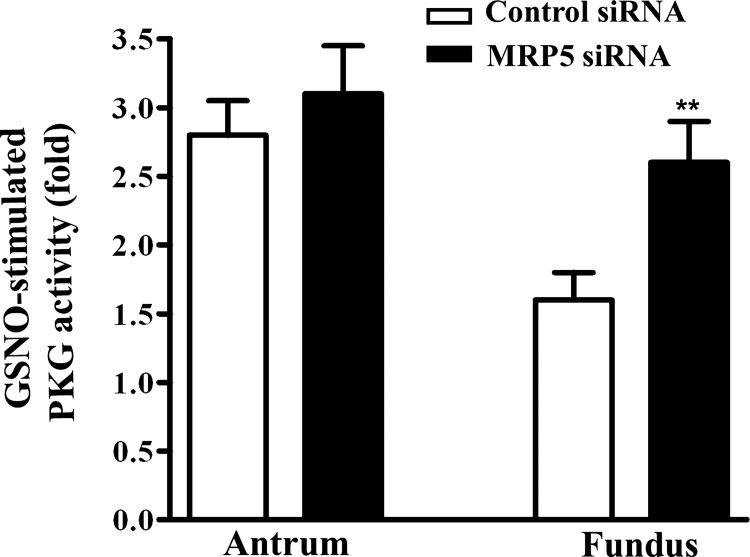
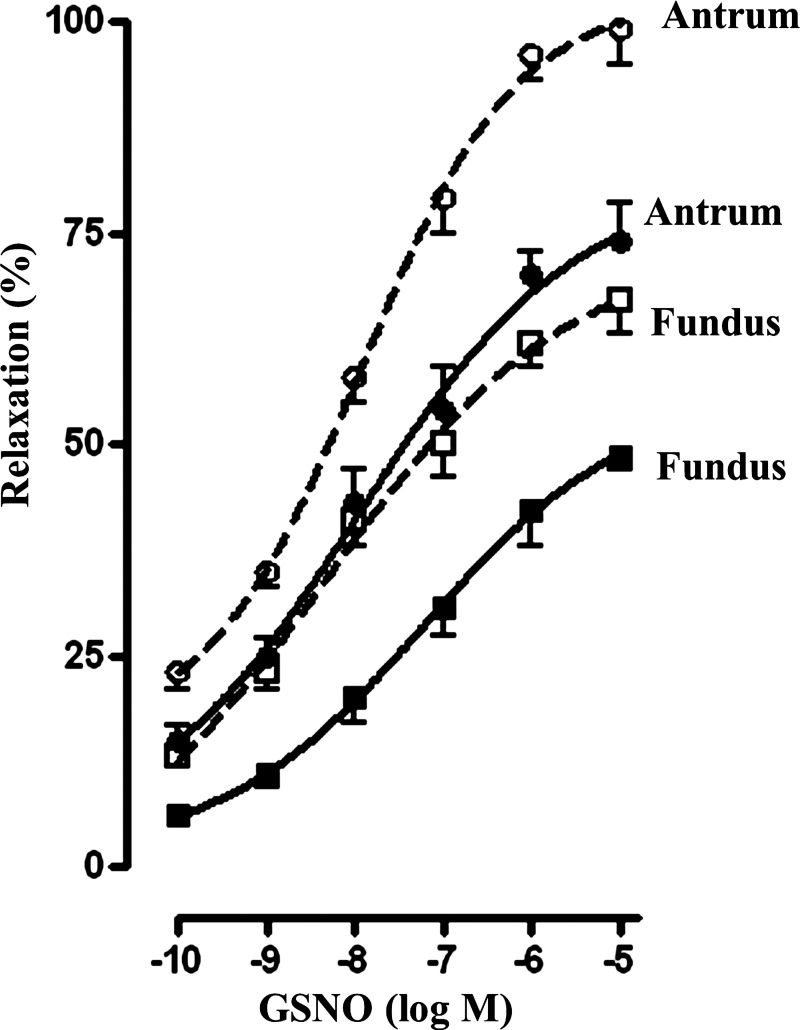
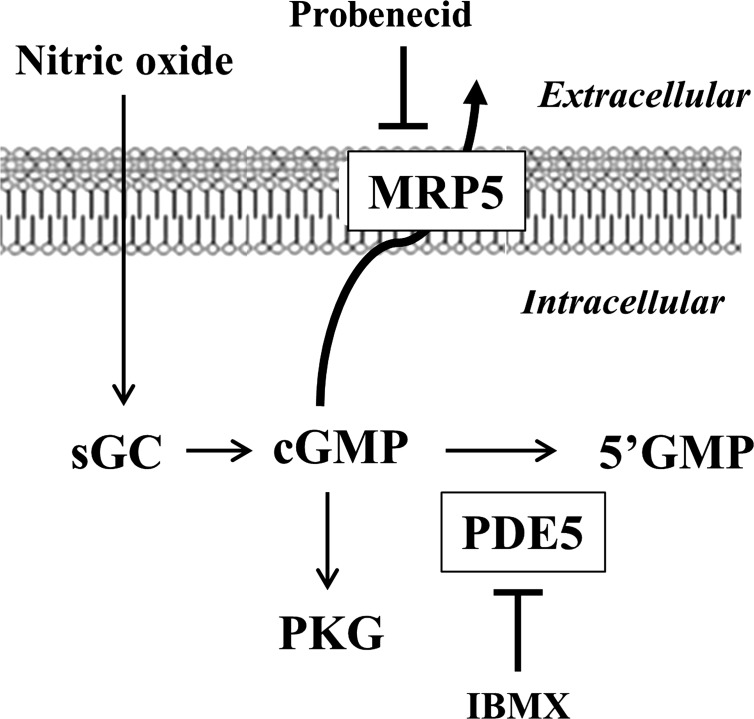
Previous studies have identified differences in the expression of proteins that regulate myosin light chain phosphorylation and contraction in tonic and phasic smooth muscle. cGMP plays a critical role in smooth muscle relaxation and is important for optimal function of phasic and tonic smooth muscle. The intracellular cGMP levels are regulated by its hydrolysis via phosphodiesterase 5 (PDE5) and efflux via novel multidrug resistance protein 5 (MRP5). In the present study we tested the hypothesis that the differences in the phasic and tonic behavior of smooth muscles may be related to differences in mechanisms that terminate cGMP signaling. Expression of PDE5 and MRP5 was significantly (more than 2-fold) higher in fundus compared with antrum. The NO donor S-nitrosoglutathione (GSNO) caused an increase in PDE5 activity and intra- and extracellular cGMP levels in both fundus and antrum. Stimulation of PDE5 activity and increase in extracellular cGMP were significantly higher in fundus, whereas increase in intracellular cGMP was significantly higher in antrum. GSNO-induced increase in extracellular cGMP was blocked in dispersed cells by the cyclic nucleotide export blocker probenecid and in cultured muscle cells by depletion of ATP or suppression of MRP5 by siRNA, providing evidence that cGMP efflux was mediated by ATP-dependent export via MRP5. Consistent with the higher expression and activity levels of PDE5 and MRP5, GSNO-induced PKG activity and muscle relaxation were significantly lower in muscle cells from fundus compared with antrum. Thus higher expression of PDE5 and MRP5 in muscle cells from fundus correlates with tonic phenotype of muscle.
Keywords: MRP5; cyclic nucleotides; phasic muscle; tonic muscle.
Figures
References
-
- Andric SA, Kostic TS, Stojilkovic SS. Contribution of multidrug resistance protein MRP5 in control of cyclic guanosine 5′-monophosphate intracellular signaling in anterior pituitary cells. Endocrinology 147: 3435–3445, 2006 - PubMed
-
- Babu GJ, Loukianov E, Loukianova T, Pyne GJ, Huke S, Osol G, Low RB, Paul RJ, Periasamy M. Loss of SM-B myosin affects muscle shortening velocity and maximal force development. Nat Cell Biol 3: 1025–1029, 2001 - PubMed
-
- Beavo JA. Cyclic nucleotide phosphodiesterases: functional implications of multiple isoforms. Physiol Rev 75: 725–748, 1995 - PubMed
-
- Bonnevier J, Fassler R, Somlyo AP, Somlyo AV, Arner A. Modulation of Ca2+ sensitivity by cyclic nucleotides in smooth muscle from protein kinase G-deficient mice. J Biol Chem 279: 5146–5151, 2004 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources