

Comprehensive analysis of the HEPN superfamily: identification of novel roles in intra-genomic conflicts, defense, pathogenesis and RNA processing
- PMID: 23768067
- PMCID: PMC3710099
- DOI: 10.1186/1745-6150-8-15
Comprehensive analysis of the HEPN superfamily: identification of novel roles in intra-genomic conflicts, defense, pathogenesis and RNA processing
Abstract
Background: The major role of enzymatic toxins that target nucleic acids in biological conflicts at all levels has become increasingly apparent thanks in large part to the advances of comparative genomics. Typically, toxins evolve rapidly hampering the identification of these proteins by sequence analysis. Here we analyze an unexpectedly widespread superfamily of toxin domains most of which possess RNase activity.
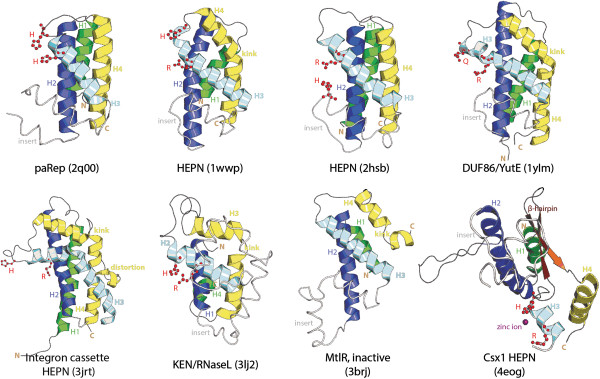
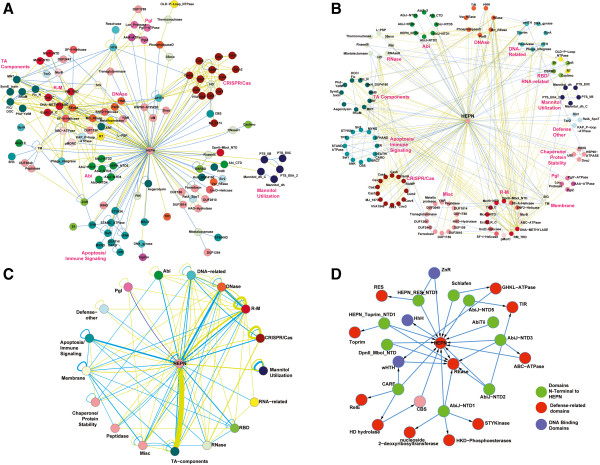
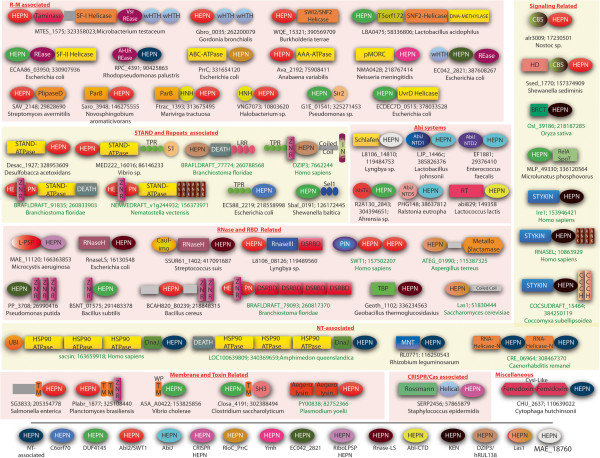
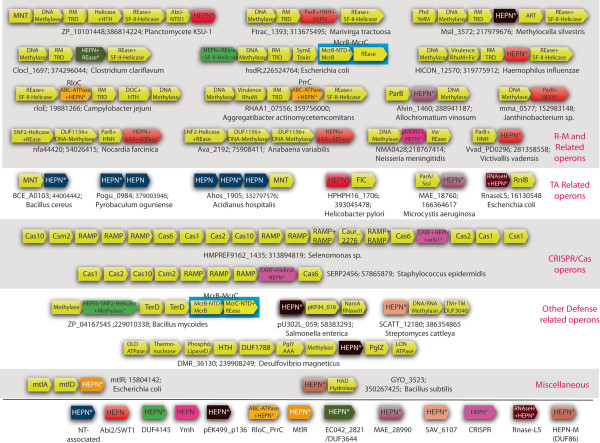
Results: The HEPN superfamily is comprised of all α-helical domains that were first identified as being associated with DNA polymerase β-type nucleotidyltransferases in prokaryotes and animal Sacsin proteins. Using sensitive sequence and structure comparison methods, we vastly extend the HEPN superfamily by identifying numerous novel families and by detecting diverged HEPN domains in several known protein families. The new HEPN families include the RNase LS and LsoA catalytic domains, KEN domains (e.g. RNaseL and Ire1) and the RNase domains of RloC and PrrC. The majority of HEPN domains contain conserved motifs that constitute a metal-independent endoRNase active site. Some HEPN domains lacking this motif probably function as non-catalytic RNA-binding domains, such as in the case of the mannitol repressor MtlR. Our analysis shows that HEPN domains function as toxins that are shared by numerous systems implicated in intra-genomic, inter-genomic and intra-organismal conflicts across the three domains of cellular life. In prokaryotes HEPN domains are essential components of numerous toxin-antitoxin (TA) and abortive infection (Abi) systems and in addition are tightly associated with many restriction-modification (R-M) and CRISPR-Cas systems, and occasionally with other defense systems such as Pgl and Ter. We present evidence of multiple modes of action of HEPN domains in these systems, which include direct attack on viral RNAs (e.g. LsoA and RNase LS) in conjunction with other RNase domains (e.g. a novel RNase H fold domain, NamA), suicidal or dormancy-inducing attack on self RNAs (RM systems and possibly CRISPR-Cas systems), and suicidal attack coupled with direct interaction with phage components (Abi systems). These findings are compatible with the hypothesis on coupling of pathogen-targeting (immunity) and self-directed (programmed cell death and dormancy induction) responses in the evolution of robust antiviral strategies. We propose that altruistic cell suicide mediated by HEPN domains and other functionally similar RNases was essential for the evolution of kin and group selection and cell cooperation. HEPN domains were repeatedly acquired by eukaryotes and incorporated into several core functions such as endonucleolytic processing of the 5.8S-25S/28S rRNA precursor (Las1), a novel ER membrane-associated RNA degradation system (C6orf70), sensing of unprocessed transcripts at the nuclear periphery (Swt1). Multiple lines of evidence suggest that, similar to prokaryotes, HEPN proteins were recruited to antiviral, antitransposon, apoptotic systems or RNA-level response to unfolded proteins (Sacsin and KEN domains) in several groups of eukaryotes.
Conclusions: Extensive sequence and structure comparisons reveal unexpectedly broad presence of the HEPN domain in an enormous variety of defense and stress response systems across the tree of life. In addition, HEPN domains have been recruited to perform essential functions, in particular in eukaryotic rRNA processing. These findings are expected to stimulate experiments that could shed light on diverse cellular processes across the three domains of life.
Reviewers: This article was reviewed by Martijn Huynen, Igor Zhulin and Nick Grishin.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
