

Widespread DNA hypomethylation at gene enhancer regions in placentas associated with early-onset pre-eclampsia
- PMID: 23770704
- PMCID: PMC3779005
- DOI: 10.1093/molehr/gat044
Widespread DNA hypomethylation at gene enhancer regions in placentas associated with early-onset pre-eclampsia
Abstract
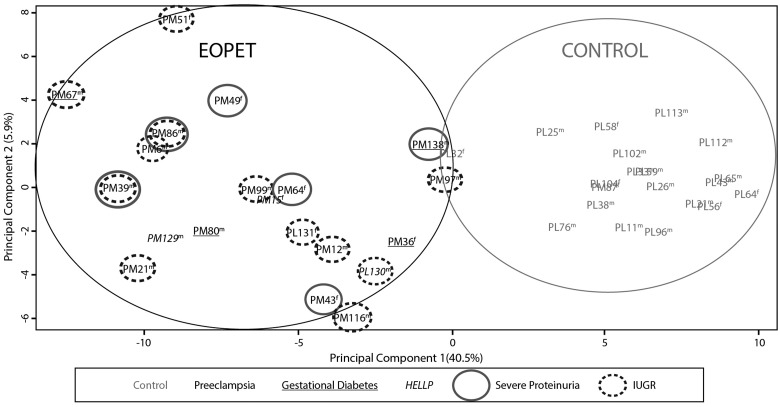
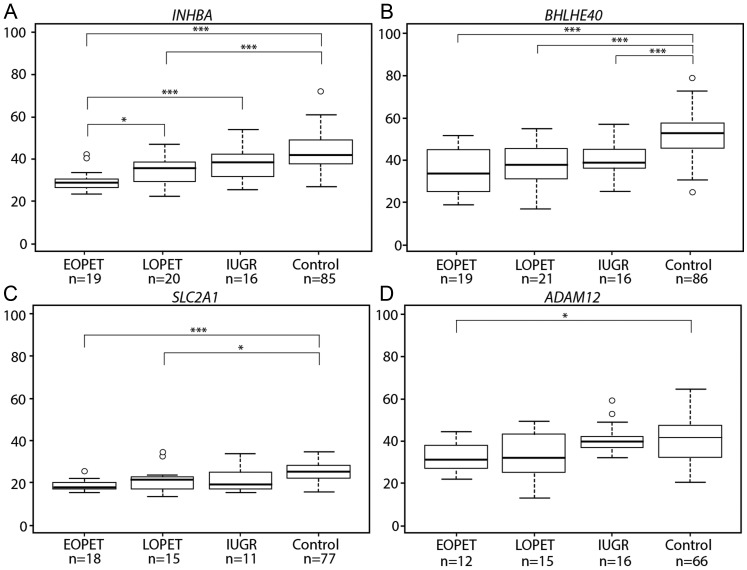
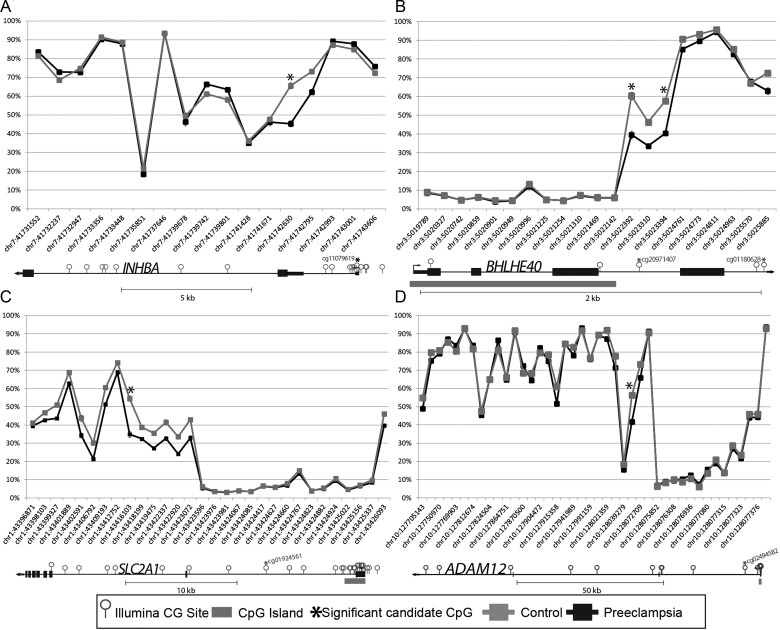
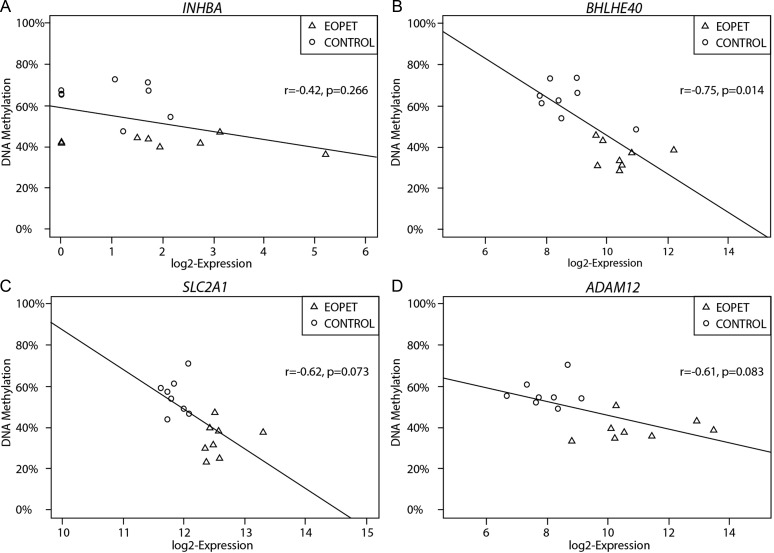
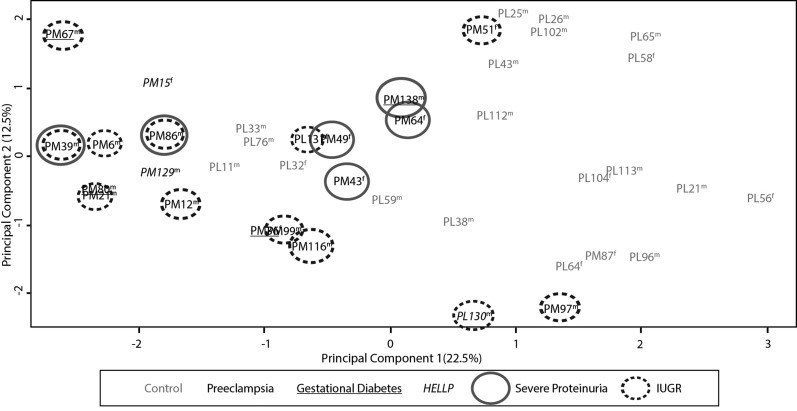
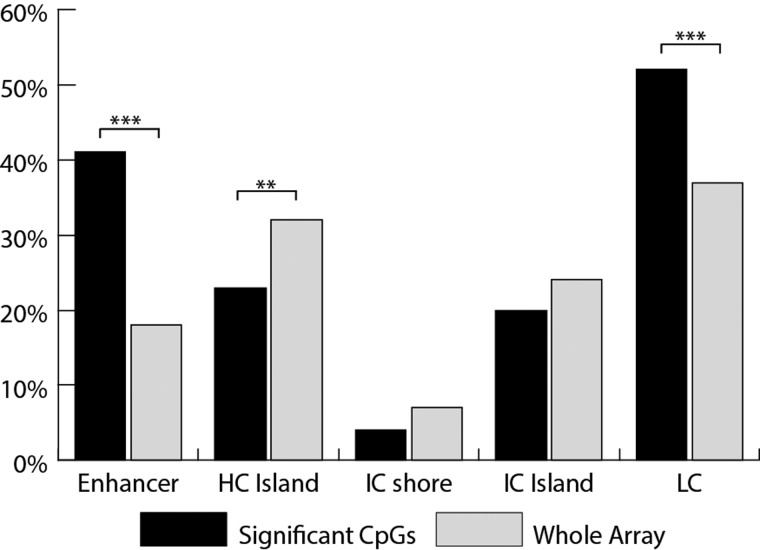
Pre-eclampsia is a serious complication of pregnancy that can affect both maternal and fetal outcomes. Early-onset pre-eclampsia (EOPET) is a severe form of pre-eclampsia that is associated with altered physiological characteristics and gene expression in the placenta. DNA methylation is a relatively stable epigenetic modification to DNA that can reflect gene expression, and can provide insight into the mechanisms underlying such expression changes. This case-control study focused on DNA methylation and gene expression of whole chorionic villi samples from 20 EOPET placentas and 20 gestational age-matched controls from pre-term births. DNA methylation was also assessed in placentas affected by late-onset pre-eclampsia (LOPET) and normotensive intrauterine growth restriction (nIUGR). The Illumina HumanMethylation450 BeadChip was used to assess DNA methylation at >480 000 cytosine-guanine dinucleotide (CpG) sites. The Illumina HT-12v4 Expression BeadChip was used to assess gene expression of >45 000 transcripts in a subset of cases and controls. DNA methylation analysis by pyrosequencing was used to follow-up the initial findings in four genes with a larger cohort of cases and controls, including nIUGR and LOPET placentas. Bioinformatic analysis was used to identify overrepresentation of gene ontology categories and transcription factor binding motifs. We identified 38 840 CpG sites with significant (false discovery rate <0.01) DNA methylation alterations in EOPET, of which 282 had >12.5% methylation difference compared with the controls. Significant sites were enriched at the enhancers and low CpG density regions of the associated genes and the majority (74.5%) of these sites were hypomethylated in EOPET. EOPET, but not associated clinical features, such as intrauterine growth restriction (IUGR), presented a distinct DNA methylation profile. CpG sites from four genes relevant to pre-eclampsia (INHBA, BHLHE40, SLC2A1 and ADAM12) showed different extent of changes in LOPET and nIUGR. Genome-wide expression in a subset of samples showed that some of the gene expression changes were negatively correlated with DNA methylation changes, particularly for genes that are responsible for angiogenesis (such as EPAS1 and FLT1). Results could be confounded by altered cell populations in abnormal placentas. Larger sample sizes are needed to fully address the possibility of sub-profiles of methylation within the EOPET cohort. Based on DNA methylation profiling, we conclude that there are widespread DNA methylation alterations in EOPET that may be associated with changes in placental function. This property may provide a useful tool for early screening of such placentas. This study identifies DNA methylation changes at many loci previously reported to have altered gene expression in EOPET placentas, as well as in novel biologically relevant genes we confirmed to be differentially expressed. These results may be useful for DNA- methylation-based non-invasive prenatal diagnosis of at-risk pregnancies.
Keywords: 450 K array; DNA methylation; placenta; pre-eclampsia.
Figures
References
-
- Alsat E, Wyplosz P, Malassine A, Guibourdenche J, Porquet D, Nessmann C, Evain-Brion D. Hypoxia impairs cell fusion and differentiation process in human cytotrophoblast, in vitro. J Cell Physiol. 1996;168:346–353. doi:10.1002/(SICI)1097-4652(199608)168:2<346::AID-JCP13>3.0.CO;2-1. - DOI - PubMed
-
- Bahado-Singh RO, Akolekar R, Mandal R, Dong E, Xia J, Kruger M, Wishart DS, Nicolaides K. First-trimester metabolomic detection of late-onset preeclampsia. Am J Obstet Gynecol. 2012;25:1840–1847. - PubMed
-
- Bailey TL, Boden M, Buske FA, Frith M, Grant CE, Clementi L, Ren J, Li WW, Noble WS. MEME SUITE: tools for motif discovery and searching. Nucleic Acids Res. 2009;37:W202–W208. doi:10.1093/nar/gkp335. - DOI - PMC - PubMed
-
- Baumann MU, Zamudio S, Illsley NP. Hypoxic upregulation of glucose transporters in BeWo choriocarcinoma cells is mediated by hypoxia-inducible factor-1. Am J Physiol Cell Physiol. 2007;293:C477–C485. doi:10.1152/ajpcell.00075.2007. - DOI - PMC - PubMed
-
- Bibikova M, Barnes B, Tsan C, Ho V, Klotzle B, Le JM, Delano D, Zhang L, Schroth GP, Gunderson KL, et al. High density DNA methylation array with single CpG site resolution. Genomics. 2011;98:288–295. doi:10.1016/j.ygeno.2011.07.007. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
