

Control of cell wall extensibility during pollen tube growth
- PMID: 23770837
- PMCID: PMC4043104
- DOI: 10.1093/mp/sst103
Control of cell wall extensibility during pollen tube growth
Abstract
In this review, we address the question of how the tip-growing pollen tube achieves its rapid rate of elongation while maintaining an intact cell wall. Although turgor is essential for growth to occur, the local expansion rate is controlled by local changes in the viscosity of the apical wall. We focus on several different structures and underlying processes that are thought to be major participants including exocytosis, the organization and activity of the actin cytoskeleton, calcium and proton physiology, and cellular energetics. We think that the actin cytoskeleton, in particular the apical cortical actin fringe, directs the flow of vesicles to the apical domain, where they fuse with the plasma membrane and contribute their contents to the expanding cell wall. While pH gradients, as generated by a proton-ATPase located on the plasma membrane along the side of the clear zone, may regulate rapid actin turnover and new polymerization in the fringe, the tip-focused calcium gradient biases secretion towards the polar axis. The recent data showing that exocytosis of new wall material precedes and predicts the process of cell elongation provide support for the idea that the intussusception of newly secreted pectin contributes to decreases in apical wall viscosity and to cell expansion. Other prime factors will be the localization and activity of the enzyme pectin methyl-esterase, and the chelation of calcium by pectic acids. Finally, we acknowledge a role for reactive oxygen species in the control of wall viscosity.
Keywords: cell expansion; cell walls; cytoskeleton dynamics; polarity; pollen development..
Figures
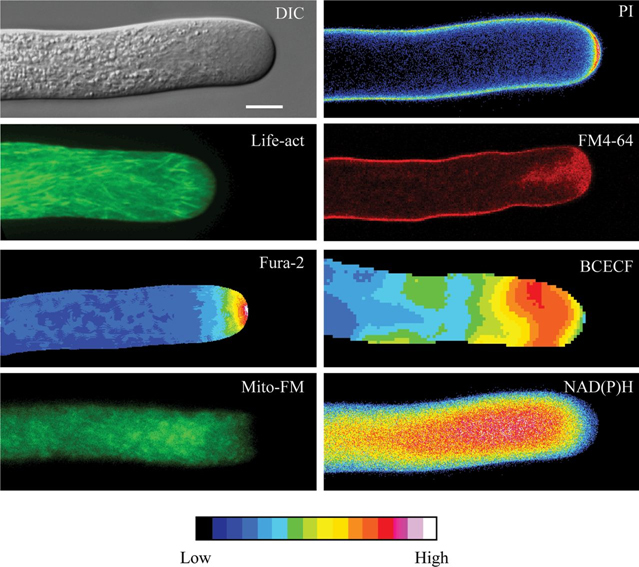
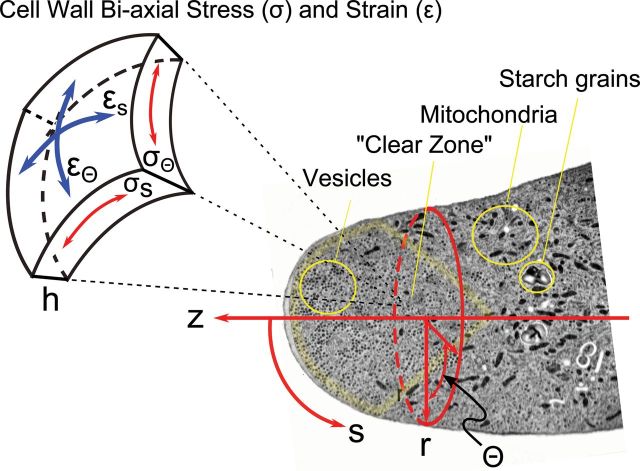
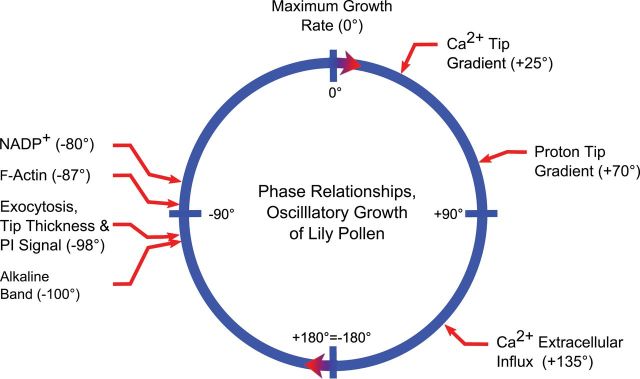
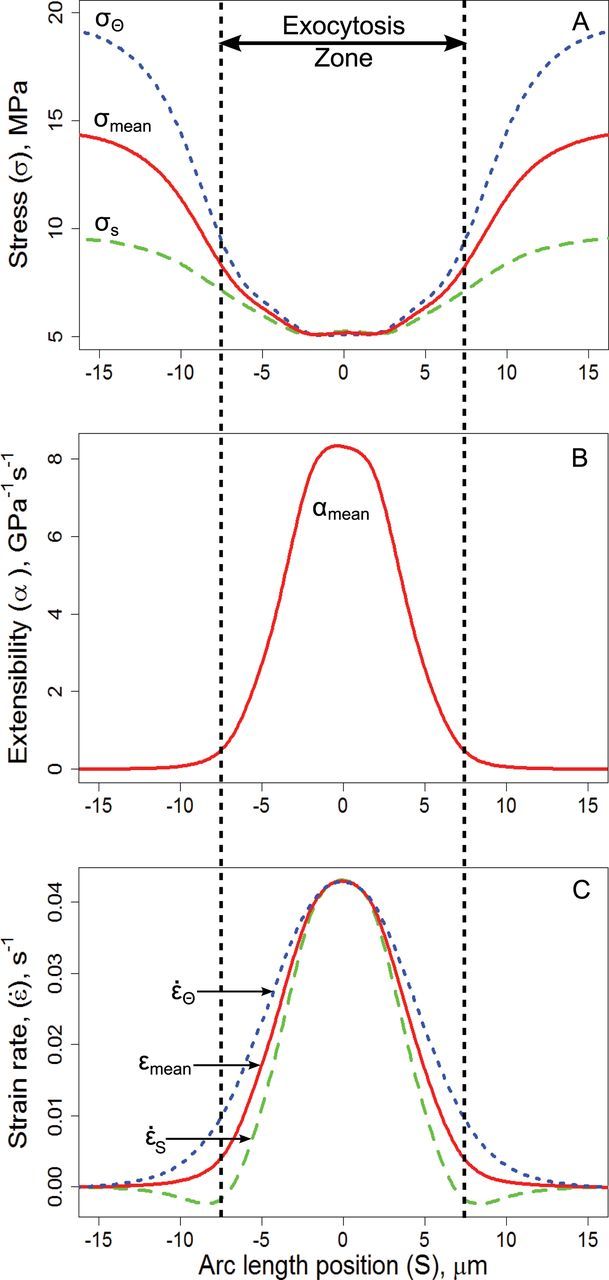
References
-
- Anderson J.R., Barnes W.S., Bedinger P. (2002). 2,6-dichlorobenzonitrile, a cellulose biosynthesis inhibitor, affects morphology and structural integrity of petunia and lily pollen tubes. J. Plant Physiol. 159, 61–67
-
- Aouar L., Chebli Y., Geitmann A. (2010). Morphogenesis of complex plant cell shapes: the mechanical role of crystalline cellulose in growing pollen tubes. Sex. Plant Reprod. 23, 15–27 - PubMed
-
- Baron-Epel O., Gharyal P.K., Schindler M. (1988). Pectins as mediators of wall porosity in soybean cells. Planta. 175, 389–395 - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
