

Gene amplification of the histone methyltransferase SETDB1 contributes to human lung tumorigenesis
- PMID: 23770855
- PMCID: PMC4031636
- DOI: 10.1038/onc.2013.239
Gene amplification of the histone methyltransferase SETDB1 contributes to human lung tumorigenesis
Abstract
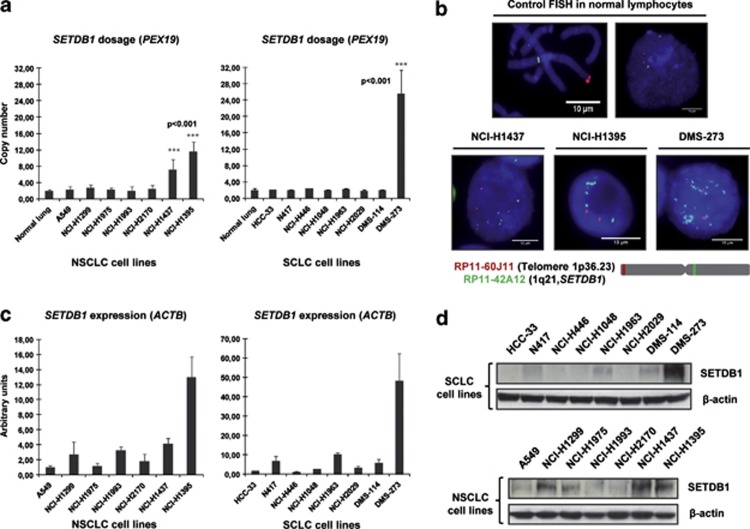
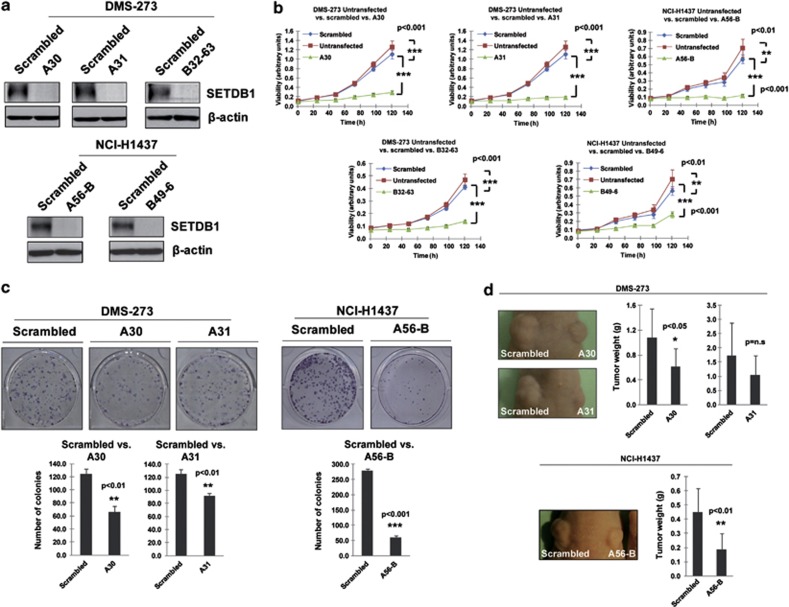
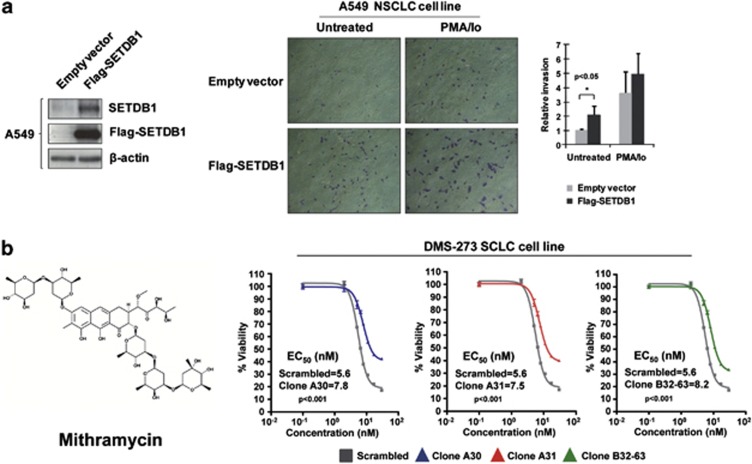
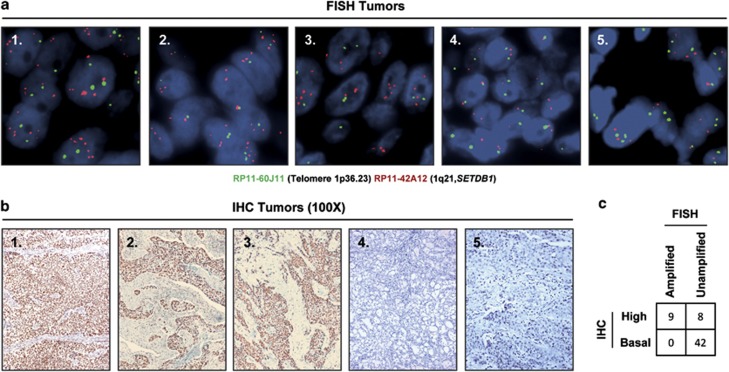
Disruption of the histone modification patterns is one of the most common features of human tumors. However, few genetic alterations in the histone modifier genes have been described in tumorigenesis. Herein we show that the histone methyltransferase SETDB1 undergoes gene amplification in non-small and small lung cancer cell lines and primary tumors. The existence of additional copies of the SETDB1 gene in these transformed cells is associated with higher levels of the corresponding mRNA and protein. From a functional standpoint, the depletion of SETDB1 expression in amplified cells reduces cancer growth in cell culture and nude mice models, whereas its overexpression increases the tumor invasiveness. The increased gene dosage of SETDB1 is also associated with enhanced sensitivity to the growth inhibitory effect mediated by the SETDB1-interfering drug mithramycin. Overall, the findings identify SETDB1 as a bona fide oncogene undergoing gene amplification-associated activation in lung cancer and suggest its potential for new therapeutic strategies.
Figures
References
-
- Berdasco M, Esteller M. Aberrant epigenetic landscape in cancer: how cellular identity goes awry. Dev Cell. 2010;19:698–711. - PubMed
-
- Füllgrabe J, Kavanagh E, Joseph B. Histone onco-modifications. Oncogene. 2011;30:3391–3403. - PubMed
-
- Fraga MF, Ballestar E, Villar-Garea A, Boix-Chornet M, Espada J, Schotta G, et al. Loss of acetylation at Lys16 and trimethylation at Lys20 of histone H4 is a common hallmark of human cancer. Nat Genet. 2005;37:391–400. - PubMed
-
- Seligson DB, Horvath S, Shi T, Yu H, Tze S, Grunstein M, et al. Global histone modification patterns predict risk of prostate cancer recurrence. Nature. 2005;435:1262–1266. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
