

Structure and function of Parkin E3 ubiquitin ligase reveals aspects of RING and HECT ligases
- PMID: 23770887
- PMCID: PMC3709503
- DOI: 10.1038/ncomms2982
Structure and function of Parkin E3 ubiquitin ligase reveals aspects of RING and HECT ligases
Abstract
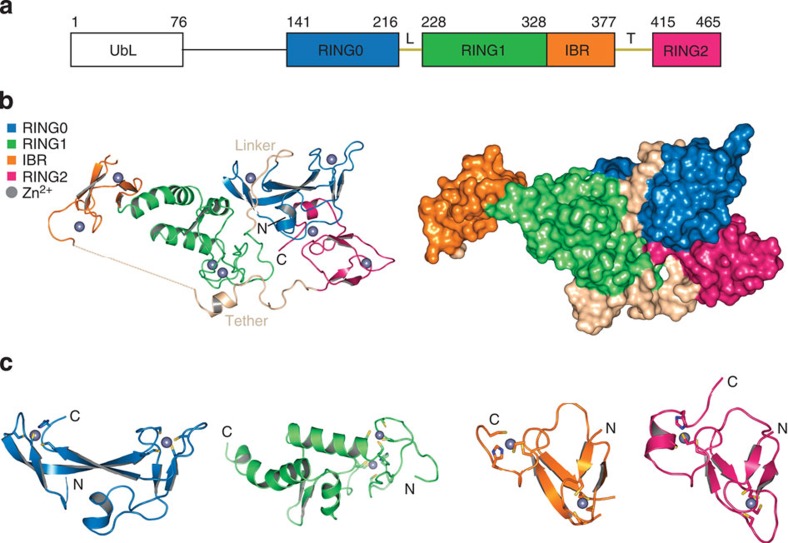
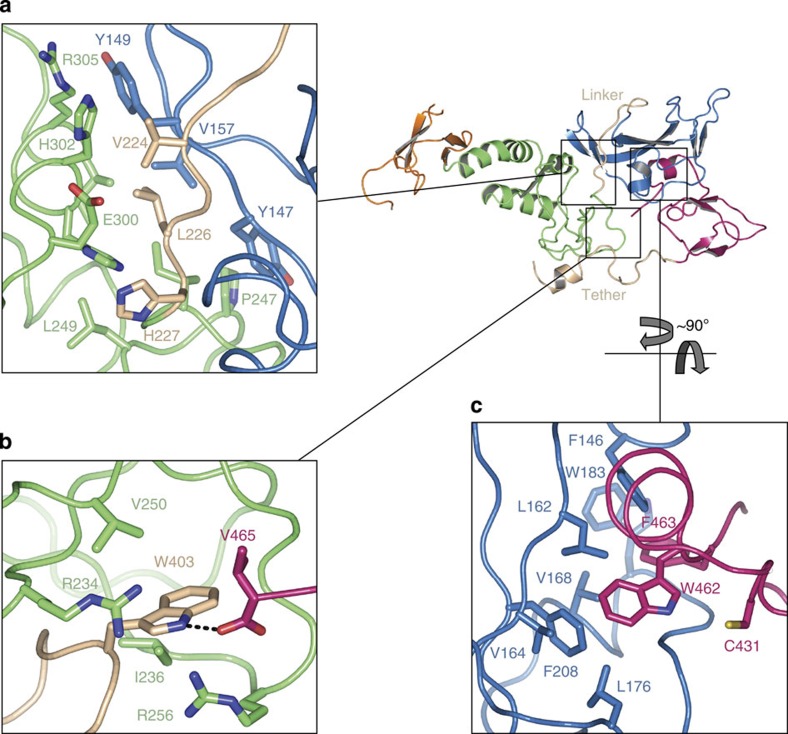
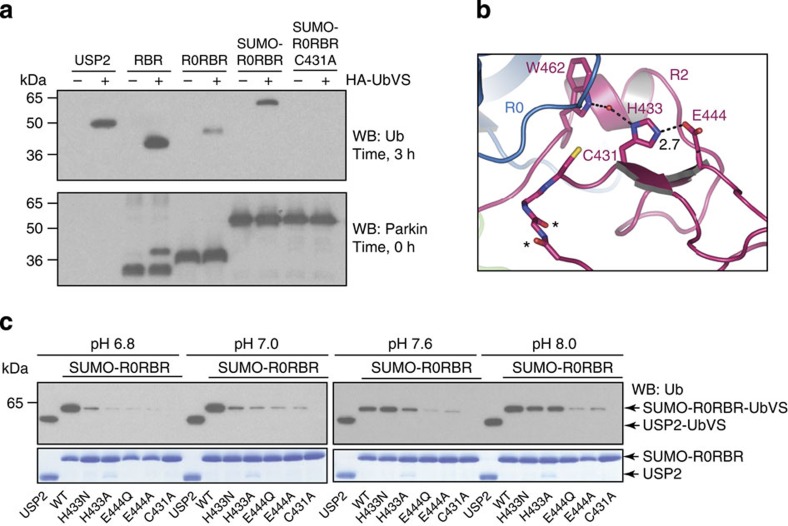
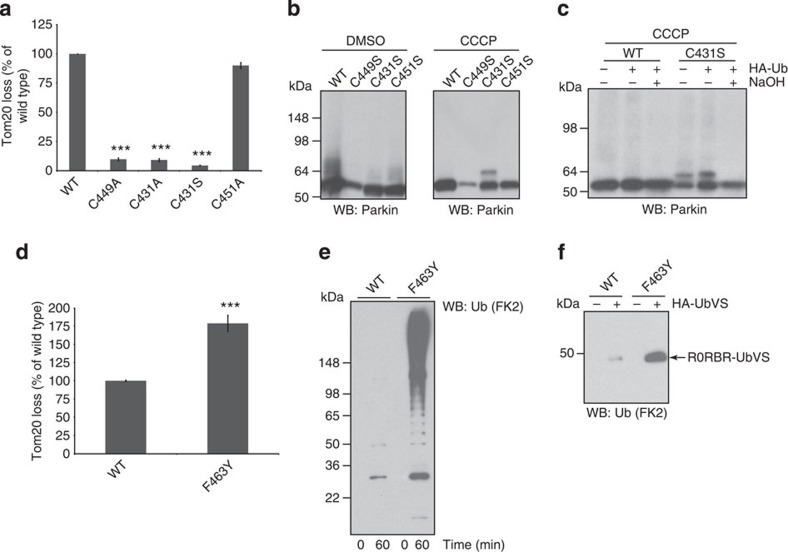
Parkin is a RING-between-RING E3 ligase that functions in the covalent attachment of ubiquitin to specific substrates, and mutations in Parkin are linked to Parkinson's disease, cancer and mycobacterial infection. The RING-between-RING family of E3 ligases are suggested to function with a canonical RING domain and a catalytic cysteine residue usually restricted to HECT E3 ligases, thus termed 'RING/HECT hybrid' enzymes. Here we present the 1.58 Å structure of Parkin-R0RBR, revealing the fold architecture for the four RING domains, and several unpredicted interfaces. Examination of the Parkin active site suggests a catalytic network consisting of C431 and H433. In cells, mutation of C431 eliminates Parkin-catalysed degradation of mitochondria, and capture of an ubiquitin oxyester confirms C431 as Parkin's cellular active site. Our data confirm that Parkin is a RING/HECT hybrid, and provide the first crystal structure of an RING-between-RING E3 ligase at atomic resolution, providing insight into this disease-related protein.
Figures
Similar articles
-
Parkin-phosphoubiquitin complex reveals cryptic ubiquitin-binding site required for RBR ligase activity.Nat Struct Mol Biol. 2017 May;24(5):475-483. doi: 10.1038/nsmb.3400. Epub 2017 Apr 17. Nat Struct Mol Biol. 2017. PMID: 28414322 Free PMC article.
-
Structure of the human Parkin ligase domain in an autoinhibited state.EMBO J. 2013 Jul 31;32(15):2099-112. doi: 10.1038/emboj.2013.125. Epub 2013 May 31. EMBO J. 2013. PMID: 23727886 Free PMC article.
-
Structure of a HOIP/E2~ubiquitin complex reveals RBR E3 ligase mechanism and regulation.Nature. 2016 Jan 28;529(7587):546-50. doi: 10.1038/nature16511. Epub 2016 Jan 20. Nature. 2016. PMID: 26789245 Free PMC article.
-
Activation of the E3 ubiquitin ligase Parkin.Biochem Soc Trans. 2015 Apr;43(2):269-74. doi: 10.1042/BST20140321. Biochem Soc Trans. 2015. PMID: 25849928 Free PMC article. Review.
-
The role of parkin in familial and sporadic Parkinson's disease.Mov Disord. 2010;25 Suppl 1(0 1):S32-9. doi: 10.1002/mds.22798. Mov Disord. 2010. PMID: 20187240 Free PMC article. Review.
Cited by
-
E3 Ubiquitin Ligases: Potential Therapeutic Targets for Skeletal Pathology and Degeneration.Stem Cells Int. 2022 Sep 27;2022:6948367. doi: 10.1155/2022/6948367. eCollection 2022. Stem Cells Int. 2022. PMID: 36203882 Free PMC article. Review.
-
Dual Function of Phosphoubiquitin in E3 Activation of Parkin.J Biol Chem. 2016 Aug 5;291(32):16879-91. doi: 10.1074/jbc.M116.728600. Epub 2016 Jun 9. J Biol Chem. 2016. PMID: 27284007 Free PMC article.
-
Proteomic Analysis of Parkin Isoforms Expression in Different Rat Brain Areas.Protein J. 2016 Oct;35(5):354-362. doi: 10.1007/s10930-016-9679-5. Protein J. 2016. PMID: 27601173
-
The Michael J. Fox Foundation for Parkinson's Research Strategy to Advance Therapeutic Development of PINK1 and Parkin.Biomolecules. 2019 Jul 24;9(8):296. doi: 10.3390/biom9080296. Biomolecules. 2019. PMID: 31344817 Free PMC article.
-
Mitophagy in Acute Kidney Injury and Kidney Repair.Cells. 2020 Feb 1;9(2):338. doi: 10.3390/cells9020338. Cells. 2020. PMID: 32024113 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
