

Sonic hedgehog and Wnt: antagonists in morphogenesis but collaborators in axon guidance
- PMID: 23772206
- PMCID: PMC3677135
- DOI: 10.3389/fncel.2013.00086
Sonic hedgehog and Wnt: antagonists in morphogenesis but collaborators in axon guidance
Abstract
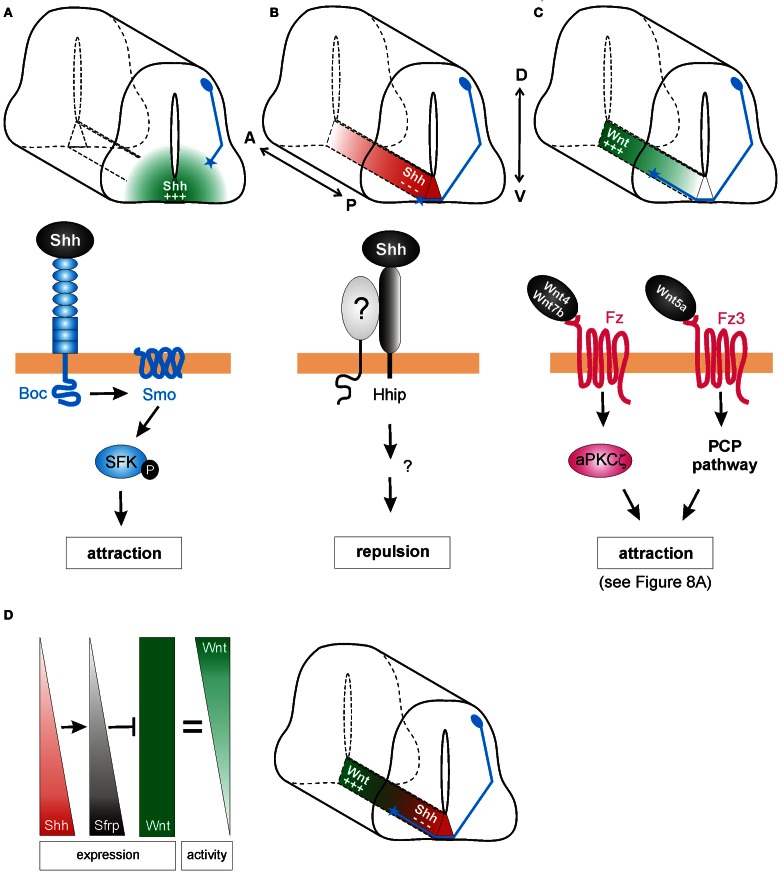
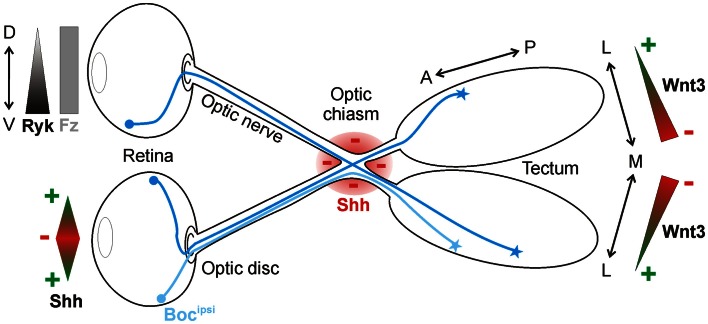
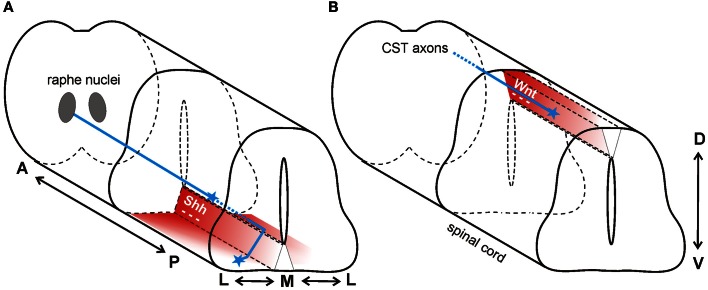
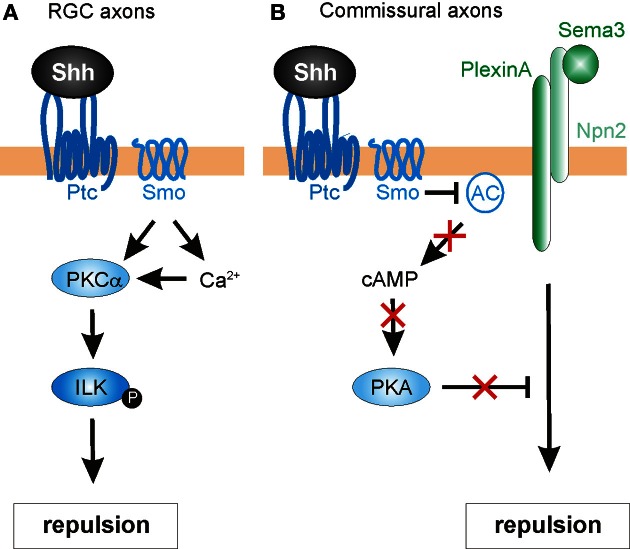
As indicated by their name, morphogens were first identified for their role in the formation of tissues early in development. Secreted from a source, they spread through the tissue to form gradients by which they affect the differentiation of precursor cells in a concentration-dependent manner. In this context, the antagonistic roles of the morphogens of the Wnt family and Sonic hedgehog (Shh) in the specification of cell types along the dorso-ventral axis of the neural tube have been studied in detail. However, more recently, morphogens have been demonstrated to act well beyond the early stages of nervous system development, as additional roles of morphogen gradients in vertebrate neural circuit formation have been identified. Both Wnt and Shh affect neural circuit formation at several stages by their influence on neurite extension, axon pathfinding and synapse formation. In this review, we will summarize the mechanisms of morphogen function during axon guidance in the vertebrate nervous system.
Keywords: Frizzled; Ryk; Smoothened; attraction; morphogen; neural circuit; repulsion; spinal cord.
Figures


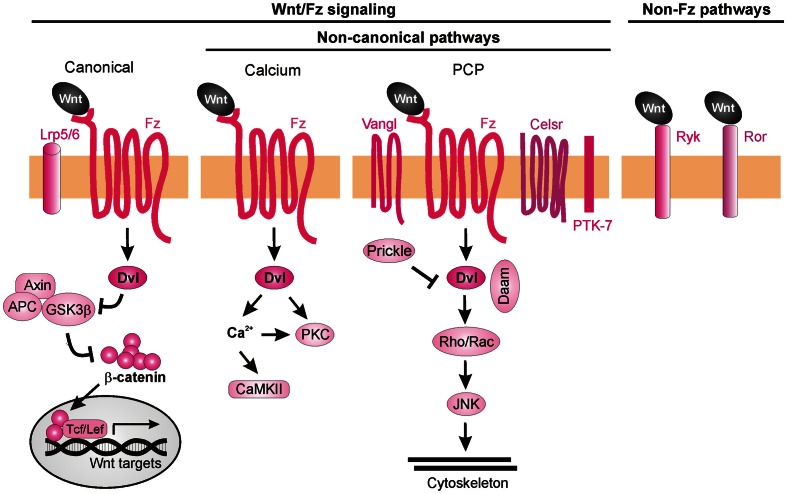
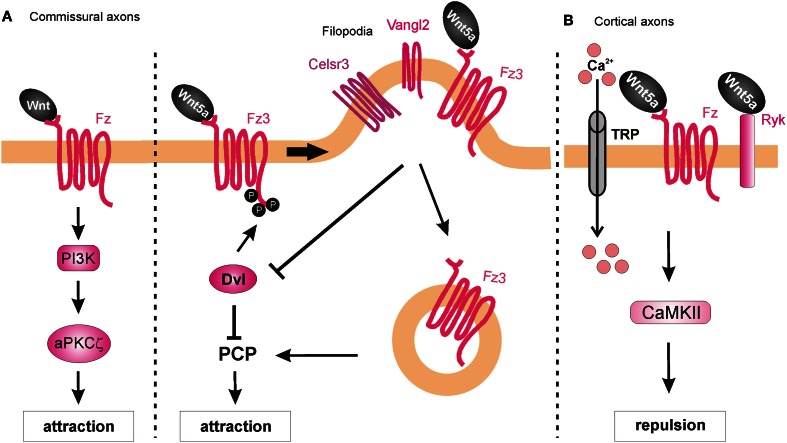
References
-
- Birgbauer E., Cowan C. A., Sretavan D. W., Henkemeyer M. (2000). Kinase independent function of EphB receptors in retinal axon pathfinding to the optic disc from dorsal but not ventral retina. Development 127, 1231–1241 - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials