

The multifaceted proprotein convertases: their unique, redundant, complementary, and opposite functions
- PMID: 23775089
- PMCID: PMC3724608
- DOI: 10.1074/jbc.R113.481549
The multifaceted proprotein convertases: their unique, redundant, complementary, and opposite functions
Abstract
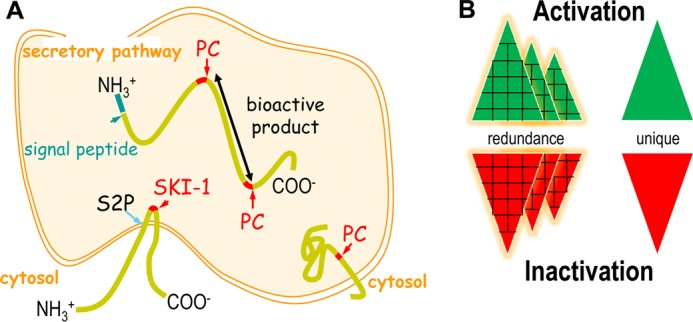
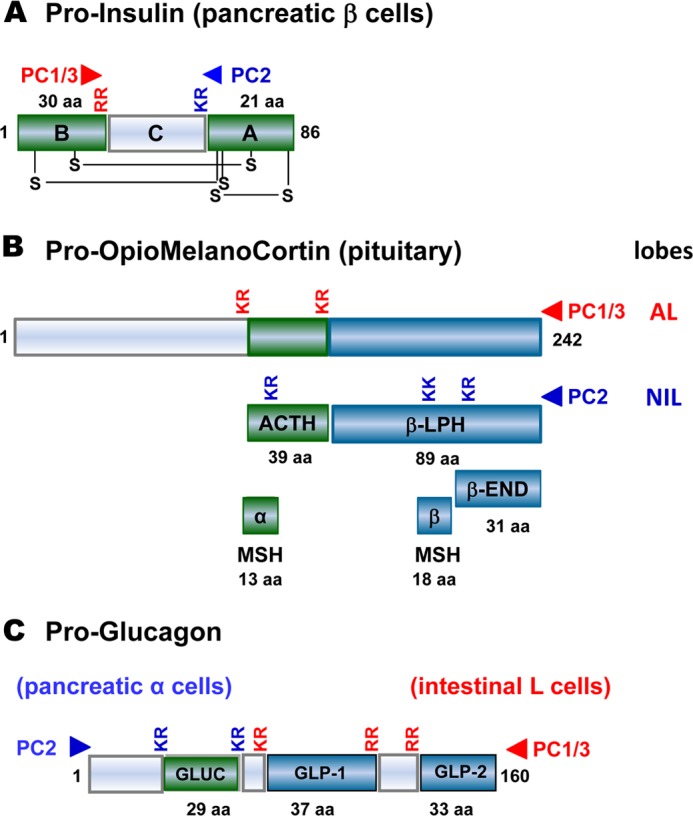
The secretory proprotein convertase (PC) family comprises nine members: PC1/3, PC2, furin, PC4, PC5/6, PACE4, PC7, SKI-1/S1P, and PCSK9. The first seven PCs cleave their substrates at single or paired basic residues, and SKI-1/S1P cleaves its substrates at non-basic residues in the Golgi. PCSK9 cleaves itself once, and the secreted inactive protease escorts specific receptors for lysosomal degradation. It regulates the levels of circulating LDL cholesterol and is considered a major therapeutic target in phase III clinical trials. In vivo, PCs exhibit unique and often essential functions during development and/or in adulthood, but certain convertases also exhibit complementary, redundant, or opposite functions.
Keywords: Cardiovascular Disease; Cell Surface Receptor; Cholesterol Metabolism; Furin-like Protease; Intracellular Processing; Low Density Lipoprotein (LDL); Precursor Processing; Proprotein Convertase; Protease; Secretory Proteins.
Figures

References
-
- Chretien M. (2011) The prohormone theory and the proprotein convertases: it is all about serendipity. Methods Mol. Biol. 768, 13–19 - PubMed
-
- Steiner D. F. (2011) On the discovery of precursor processing. Methods Mol. Biol. 768, 3–11 - PubMed
-
- Seidah N. G., Prat A. (2012) The biology and therapeutic targeting of the proprotein convertases. Nat. Rev. Drug Discov. 11, 367–383 - PubMed
-
- Steiner D. F. (1998) The proprotein convertases. Curr. Opin. Chem. Biol. 2, 31–39 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
