

Arabidopsis MSI1 connects LHP1 to PRC2 complexes
- PMID: 23778966
- PMCID: PMC3715863
- DOI: 10.1038/emboj.2013.145
Arabidopsis MSI1 connects LHP1 to PRC2 complexes
Abstract
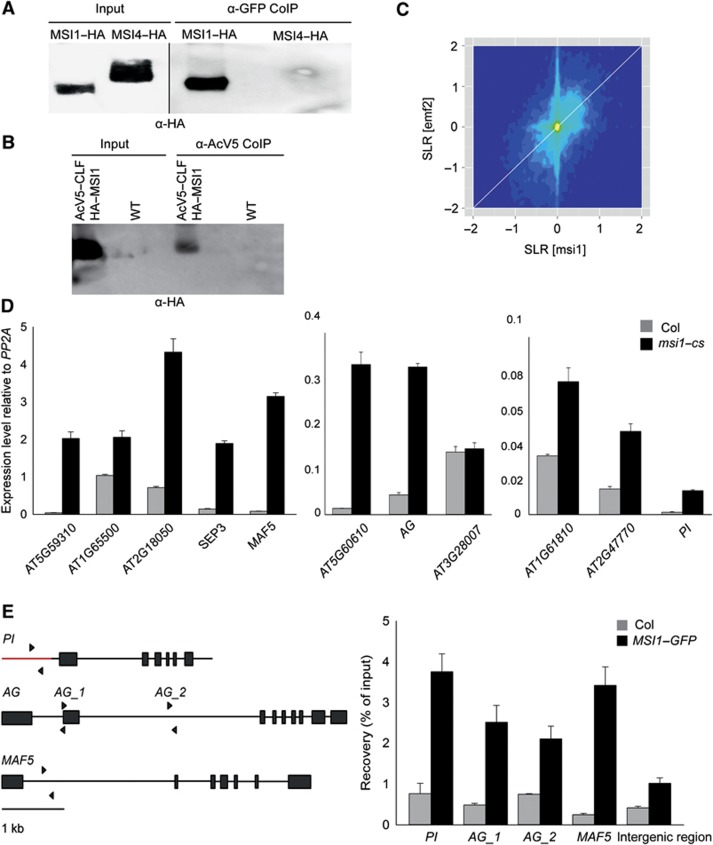
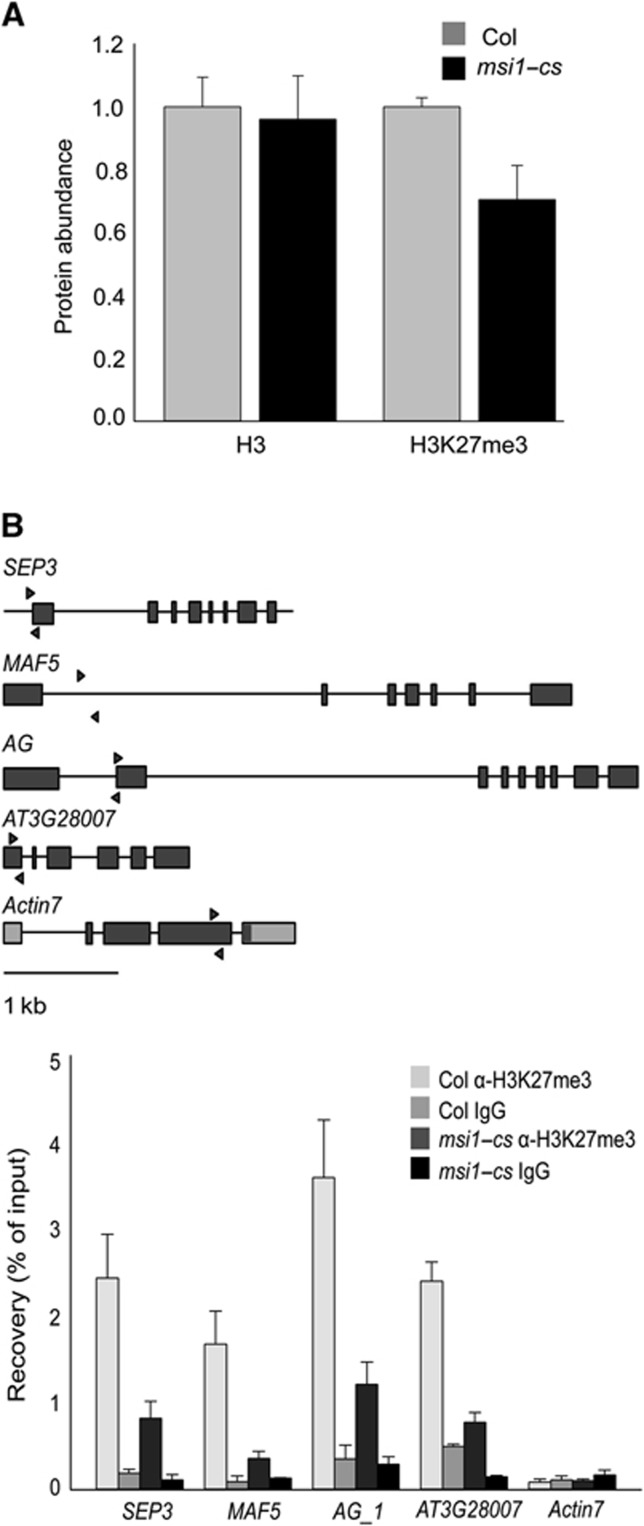
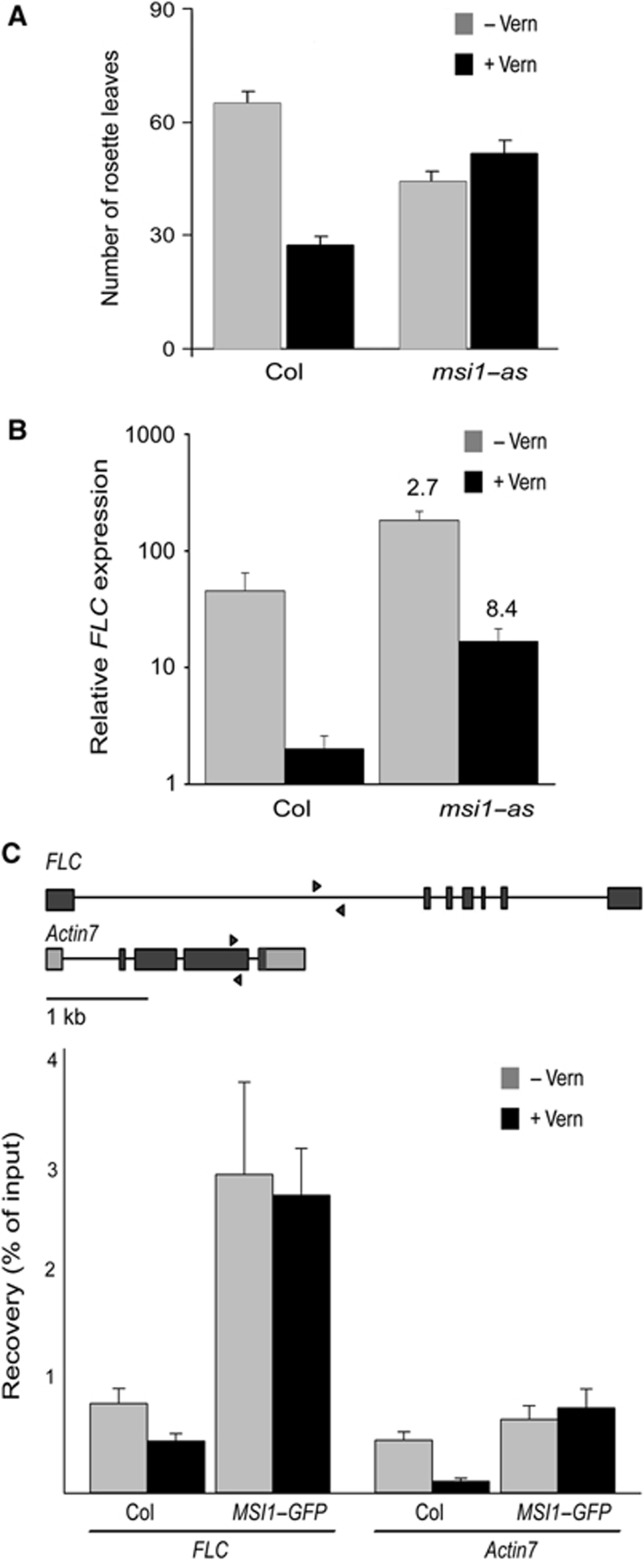
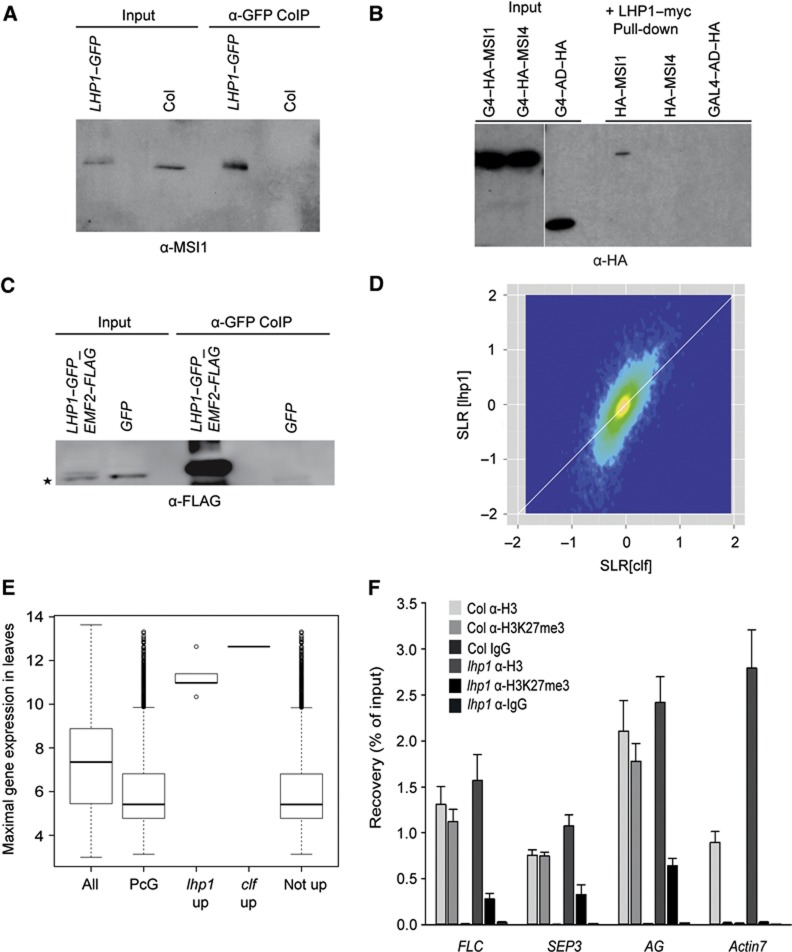
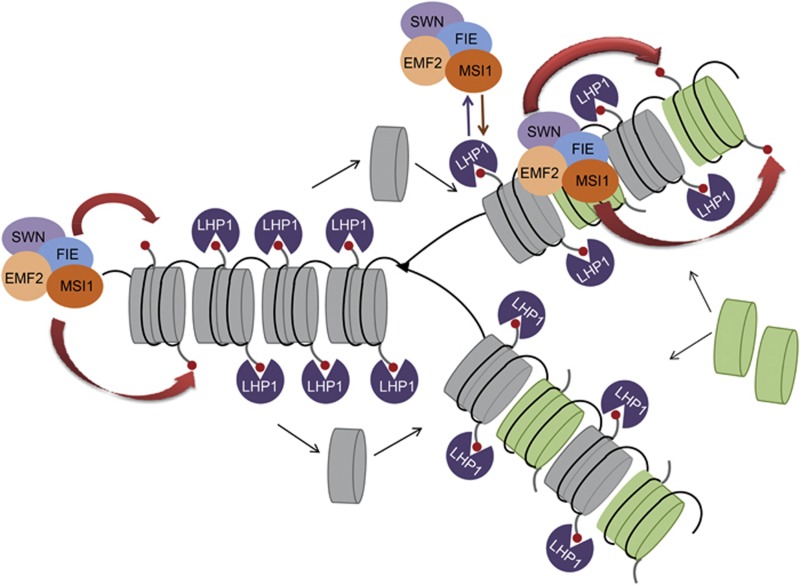
Polycomb group (PcG) proteins form essential epigenetic memory systems for controlling gene expression during development in plants and animals. However, the mechanism of plant PcG protein functions remains poorly understood. Here, we probed the composition and function of plant Polycomb repressive complex 2 (PRC2). This work established the fact that all known plant PRC2 complexes contain MSI1, a homologue of Drosophila p55. While p55 is not essential for the in vitro enzymatic activity of PRC2, plant MSI1 was required for the functions of the EMBRYONIC FLOWER and the VERNALIZATION PRC2 complexes including trimethylation of histone H3 Lys27 (H3K27) at the target chromatin, as well as gene repression and establishment of competence to flower. We found that MSI1 serves to link PRC2 to LIKE HETEROCHROMATIN PROTEIN 1 (LHP1), a protein that binds H3K27me3 in vitro and in vivo and is required for a functional plant PcG system. The LHP1-MSI1 interaction forms a positive feedback loop to recruit PRC2 to chromatin that carries H3K27me3. Consequently, this can provide a mechanism for the faithful inheritance of local epigenetic information through replication.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
Similar articles
-
The domesticated transposase ALP2 mediates formation of a novel Polycomb protein complex by direct interaction with MSI1, a core subunit of Polycomb Repressive Complex 2 (PRC2).PLoS Genet. 2020 May 28;16(5):e1008681. doi: 10.1371/journal.pgen.1008681. eCollection 2020 May. PLoS Genet. 2020. PMID: 32463832 Free PMC article.
-
Kicking against the PRCs - A Domesticated Transposase Antagonises Silencing Mediated by Polycomb Group Proteins and Is an Accessory Component of Polycomb Repressive Complex 2.PLoS Genet. 2015 Dec 7;11(12):e1005660. doi: 10.1371/journal.pgen.1005660. eCollection 2015 Dec. PLoS Genet. 2015. PMID: 26642436 Free PMC article.
-
The chromodomain of LIKE HETEROCHROMATIN PROTEIN 1 is essential for H3K27me3 binding and function during Arabidopsis development.PLoS One. 2009;4(4):e5335. doi: 10.1371/journal.pone.0005335. Epub 2009 Apr 28. PLoS One. 2009. PMID: 19399177 Free PMC article.
-
Polycomb group and trithorax group proteins in Arabidopsis.Biochim Biophys Acta. 2007 May-Jun;1769(5-6):375-82. doi: 10.1016/j.bbaexp.2007.01.010. Epub 2007 Feb 7. Biochim Biophys Acta. 2007. PMID: 17363079 Review.
-
Diversity of Polycomb group complexes in plants: same rules, different players?Trends Genet. 2009 Sep;25(9):414-23. doi: 10.1016/j.tig.2009.07.002. Epub 2009 Aug 27. Trends Genet. 2009. PMID: 19716619 Review.
Cited by
-
GmBTB/POZ promotes the ubiquitination and degradation of LHP1 to regulate the response of soybean to Phytophthora sojae.Commun Biol. 2021 Mar 19;4(1):372. doi: 10.1038/s42003-021-01907-7. Commun Biol. 2021. PMID: 33742112 Free PMC article.
-
The Harbinger transposon-derived gene PANDA epigenetically coordinates panicle number and grain size in rice.Plant Biotechnol J. 2022 Jun;20(6):1154-1166. doi: 10.1111/pbi.13799. Epub 2022 Mar 16. Plant Biotechnol J. 2022. PMID: 35239255 Free PMC article.
-
Arabidopsis MSI1 functions in photoperiodic flowering time control.Front Plant Sci. 2014 Mar 7;5:77. doi: 10.3389/fpls.2014.00077. eCollection 2014. Front Plant Sci. 2014. PMID: 24639681 Free PMC article.
-
Dynamic regulation and function of histone monoubiquitination in plants.Front Plant Sci. 2014 Mar 13;5:83. doi: 10.3389/fpls.2014.00083. eCollection 2014. Front Plant Sci. 2014. PMID: 24659991 Free PMC article. Review.
-
CENTRORADIALIS Interacts with FLOWERING LOCUS T-Like Genes to Control Floret Development and Grain Number.Plant Physiol. 2019 Jun;180(2):1013-1030. doi: 10.1104/pp.18.01454. Epub 2019 Apr 19. Plant Physiol. 2019. PMID: 31004004 Free PMC article.
References
-
- Alexandre C, Möller-Steinbach Y, Schönrock N, Gruissem W, Hennig L (2009) Arabidopsis MSI1 is required for negative regulation of the response to drought stress. Mol Plant 2: 675–687 - PubMed
-
- Aranda B, Achuthan P, Alam-Faruque Y, Armean I, Bridge A, Derow C, Feuermann M, Ghanbarian AT, Kerrien S, Khadake J, Kerssemakers J, Leroy C, Menden M, Michaut M, Montecchi-Palazzi L, Neuhauser SN, Orchard S, Perreau V, Roechert B, van Eijk K et al. (2010) The IntAct molecular interaction database in 2010. Nucleic Acids Res 38: D525–D531 - PMC - PubMed
-
- Ausin I, Alonso-Blanco C, Jarillo JA, Ruiz-Garcia L, Martinez-Zapater JM (2004) Regulation of flowering time by FVE, a Retinoblastoma-associated protein. Nat Genet 36: 162–166 - PubMed
-
- Baerenfaller K, Hirsch-Hoffmann M, Svozil J, Hull R, Russenberger D, Bischof S, Lu Q, Gruissem W, Baginsky S (2011) pep2pro: a new tool for comprehensive proteome data analysis to reveal information about organ-specific proteomes in Arabidopsis thaliana. Integr Biol 3: 225–237 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
