

Review
doi: 10.1038/nrm3609.
Epub 2013 Jun 19.
Functions of cofilin in cell locomotion and invasion
Affiliations
- PMID: 23778968
- PMCID: PMC3878614
- DOI: 10.1038/nrm3609
Item in Clipboard
Review
Functions of cofilin in cell locomotion and invasion
Nat Rev Mol Cell Biol.
2013 Jul.
Abstract
Recently, a consensus has emerged that cofilin severing activity can generate free actin filament ends that are accessible for F-actin polymerization and depolymerization without changing the rate of G-actin association and dissociation at either filament end. The structural basis of actin filament severing by cofilin is now better understood. These results have been integrated with recently discovered mechanisms for cofilin activation in migrating cells, which led to new models for cofilin function that provide insights into how cofilin regulation determines the temporal and spatial control of cell behaviour.
Figures
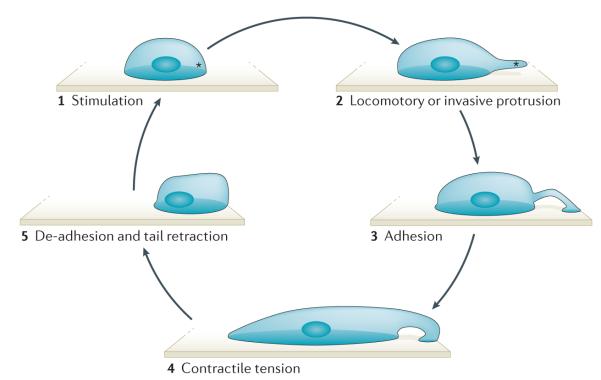
An external or internal signal stimulates the local and asymmetric polymerization of actin (depicted by an asterisk) (1). Polymerizing actin pushes against the cell membrane to form a locomotory or invasive protrusion (2). The attachment of the protrusion to the extracellular matrix initiates adhesion formation at the leading edge and signals to the contractile machinery of the cell to initiate contraction (3). Contractile tension distorts the cell into an elongated shape (4). Release of the rear adhesion allows tail retraction and locomotion of the cell (5).
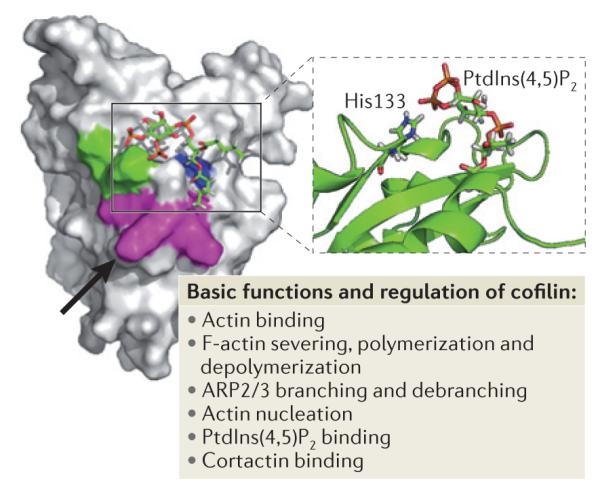
Key amino acids involved in the regulation of cofilin function are Asp122 and His133. The black arrow points to Asp122 located underneath Leu126 (in magenta). Asp122 regulates binding of cofilin to phosphatidylinositol-4,5-bisphophate (PtdIns(4,5)P2). A cofilin mutant in which Asp122 has been replaced with Ala has increased binding affinity for PtdIns (4,5)P2 and this results in the inhibition of actin polymerization, cell protrusion and migration. His133 decreases cofilin affinity for PtdIns(4,5)P2 when deprotonated. Substitution of His133 with Ala in cofilin causes loss of H+ binding and increases steady-state actin polymerization and cell protrusion. Lys132 (which is involved in actin binding) and His133 are shown in green, Lys125, Lys126, and Lys127 (which are alternative residues for PtdIns(4,5)P2 binding) are shown in magenta and Phe15 and Leu99 are shown in blue. The figure is modified, with permission, from REF. 102 © (2008) Rockefeller University Press.
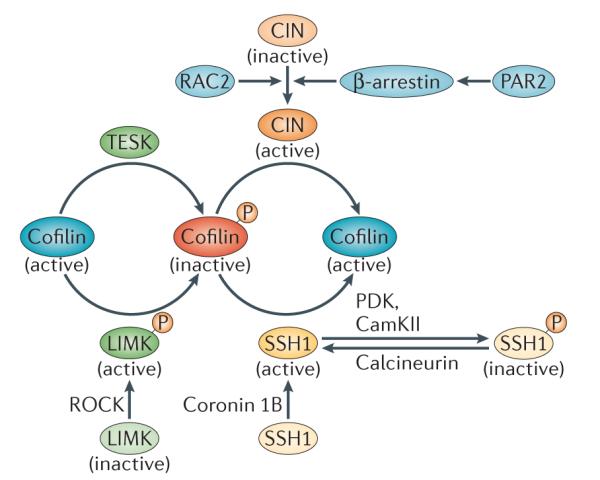
Cofilin phosphorylation at Ser3 is driven by LIM-domain kinase (LIMK) and TES kinase (TESK). LIMK is activated by ROH-associated protein kinase (ROCK). Dephosphorylation of cofilin at Ser3 is mediated by the phosphatases chronophin (CIN) and slinghshot 1 (SSH1).The localization and activation of SSH1 is mediated by coronin 1B. Phosphorylation of SSH1 by Ca2+/calmodulin-dependent protein kinase II (CaMKII) and phosphoinositide-dependent protein kinase (PDK) results in its inactivation. Calcineurin activates SSH1 by dephosphorylating it. CIN is activated by its interaction with β-arrestin, which is activated by protease-activated receptor 2 (PAR2). In neutrophils, RAC2 has been shown to induce cofilin activation in a CIN-dependent manner.
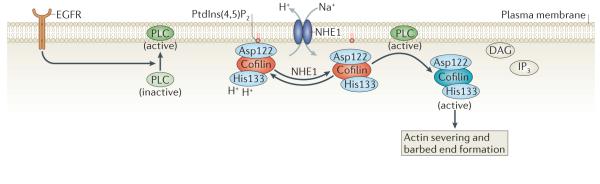
Inactive cofilin is bound to phosphatidylinositol-4,5-bisphosphate (PtdIns(4,5)P2) at the plasma membrane through its Asp122 residue. Na+–H+ exchanger 1 (NHE1) can increase the intracellular pH by exchanging intracellular H+ for extracellular Na+. This causes the deprotonation of cofilin at His133, which alters its binding affinity for PtdIns(4,5)P2. An increased pH facilitates phospholipase C (PLC)-mediated hydrolysis of PtdIns(4,5)P2 to diacylglycerol (DAG) and inositol-1,4,5,-trisphosphate (IP3) and the release of cofilin from PtdIns(4,5)P2. Epidermal growth factor receptor (EGFR) stimulates PLC activity, which in turn increases cofilin activation. See BOX 1 for methods describing how to study cofilin mobility as well as cofilin binding to the plasma membrane and F-actin.
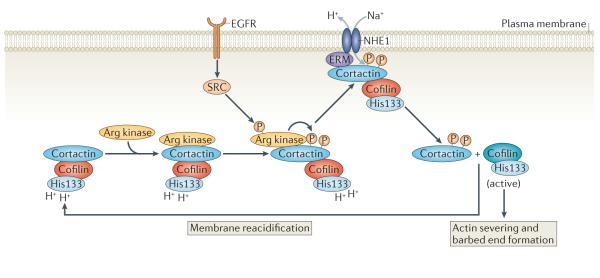
Tyr phosphorylation of cortactin by Arg kinase, which is activated downstream of epidermal growth factor receptor (EGFR), regulates the interaction between the Na+–H+ exchanger 1 (NHE1) and cortactin. The recruitment of phosphorylated cortactin to NHE1 may be mediated by ERM (ezrin–radixin–moesin) proteins. NHE1 increases the intracellular pH, which induces the release of cortactin-bound cofilin.
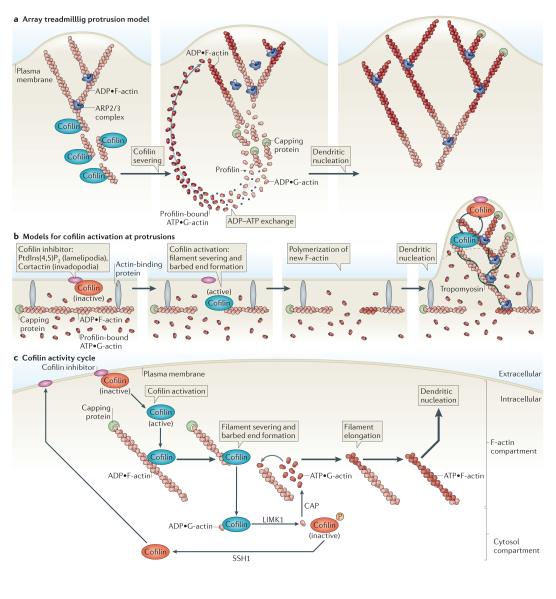
a ∣ Array treadmilling protrusion model. Cofilin severs and depolymerizes actin filaments at the base of the lamellipodium, thereby supplying G-actin monomers for steady-state actin polymerization in conditions of G-actin depletion. Dendritic nucleation is mediated by the actin-related protein 2/3 (ARP2/3) complex. b ∣ Models for cofilin activation at protrusions. Release of cofilin from its inhibitor (phosphatidylinositol-4,5-bisphosphate (PtdIns(4,5)P2) at lamellipodia or cortactin at invadopodia) at the plasma membrane increases severing of actin filaments, generating free barbed ends that define the sites of dendritic nucleation by the ARP2/3 complex. G-actin monomers are supplied from an abundant pre-existing G-actin pool. The ARP2/3 complex mediates dendritic nucleation. Tropomyosin limits cofilin action, as it inhibits binding of cofilin to F-actin. This confines cofilin severing to the tip of protrusions (dotted black line), where cofilin continues its cycles of activation and deactivation due to the local cofilin activity cycle. c ∣ Cofilin activity cycle. Following its dephosphorylation or its release from inhibitory binding partners, activated cofilin binds to and severs actin filaments, generating barbed ends that are used for actin polymerization. LIM kinase 1 (LIMK1) and cyclase-associated protein (CAP) accelerate the dissociation of ADP•G-actin–cofilin complexes, and this increases the concentration of ATP•G-actin monomers available for filament elongation at the newly formed barbed ends. The capping of new barbed ends by actin interacting protein 1 (AIP1) (not shown) promotes cofilin-dependent filament disassembly. LIMK1 phosphorylates cofilin and thus inactivates it, leading to the release of bound G-actin. Upon the activation of slingshot 1 (SSH1) , cofilin is dephosphorylated and recycled to the inhibitory compartment. Figures are not drawn to scale. See BOX 1 for methods describing how to study cofilin activation.
References
-
- Charras G, Paluch E. Blebs lead the way: how to migrate without lamellipodia. Nature Rev. Mol. Cell Biol. 2008;9:730–736. - PubMed
-
- Daubon T, Rochelle T, Bourmeyster N, Genot E. Invadopodia and rolling-type motility are specific features of highly invasive p190bcr-abl leukemic cells. Eur. J. Cell Biol. 2012;91:978–987. - PubMed
-
- Nurnberg A, Kitzing T, Grosse R. Nucleating actin for invasion. Nature Rev. Cancer. 2011;11:177–187. - PubMed
-
- Ridley AJ. Life at the leading edge. Cell. 2011;145:1012–1022. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
