

Inhibition of TGF-β signaling at the nuclear envelope: characterization of interactions between MAN1, Smad2 and Smad3, and PPM1A
- PMID: 23779087
- PMCID: PMC3843637
- DOI: 10.1126/scisignal.2003411
Inhibition of TGF-β signaling at the nuclear envelope: characterization of interactions between MAN1, Smad2 and Smad3, and PPM1A
Abstract
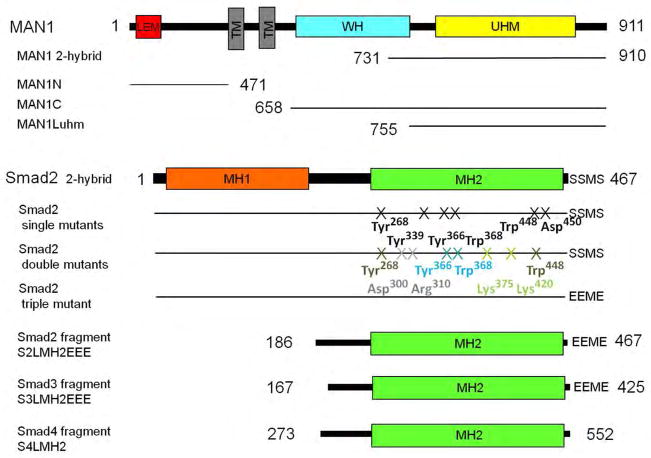
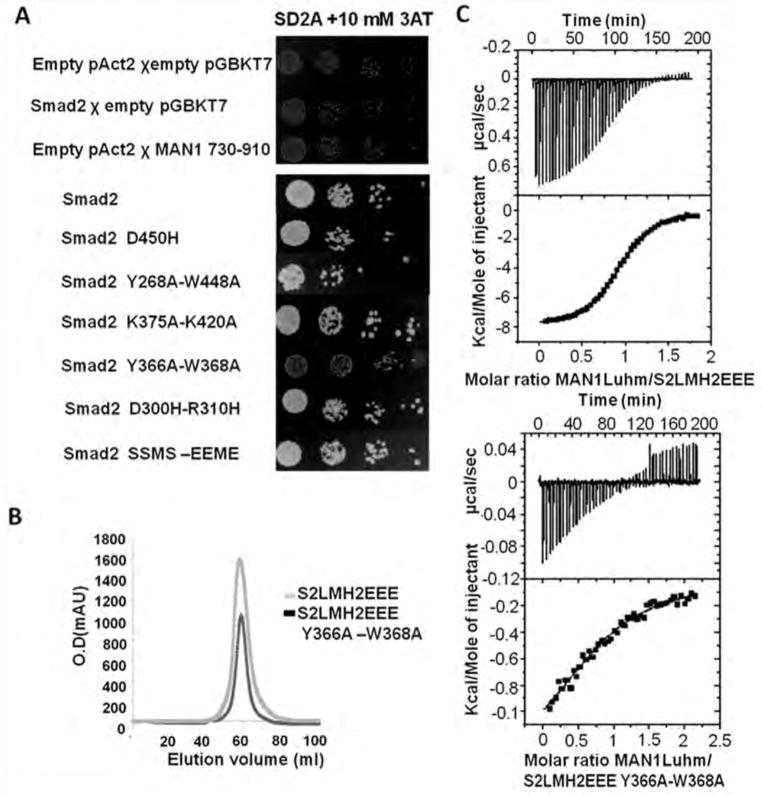
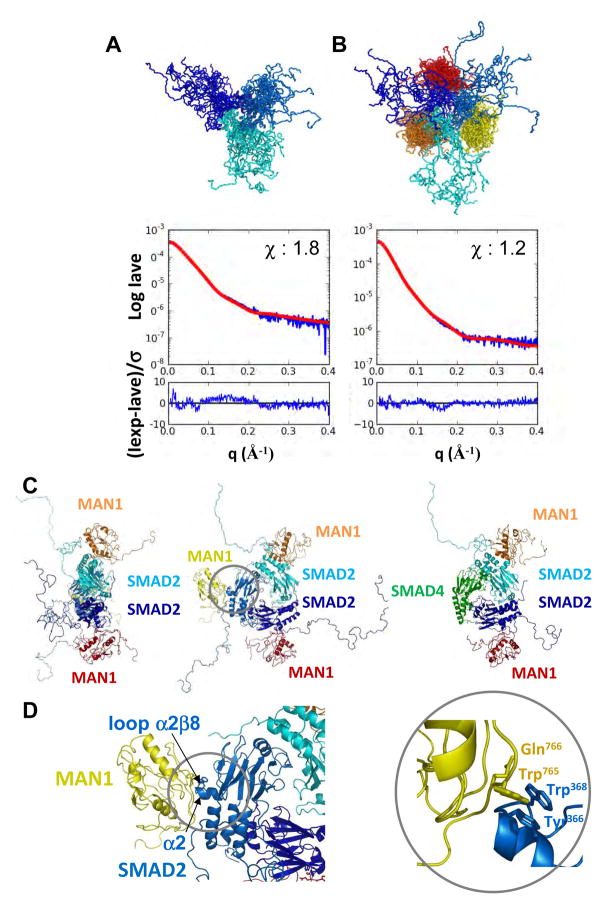
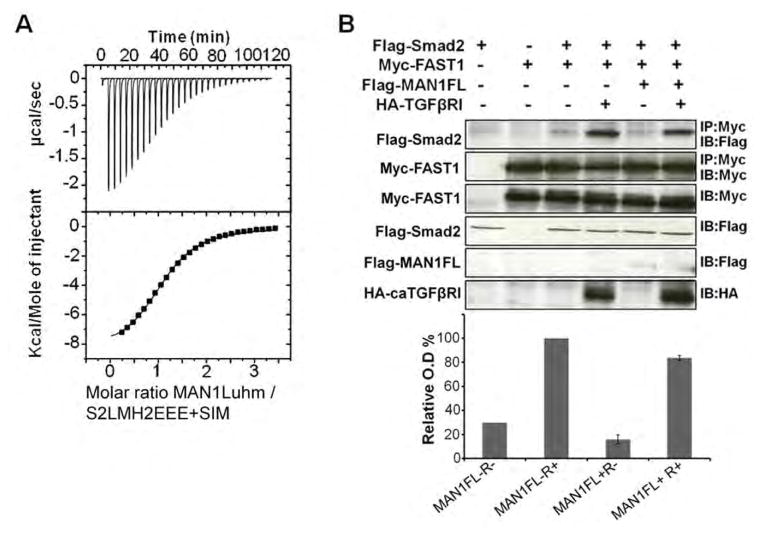
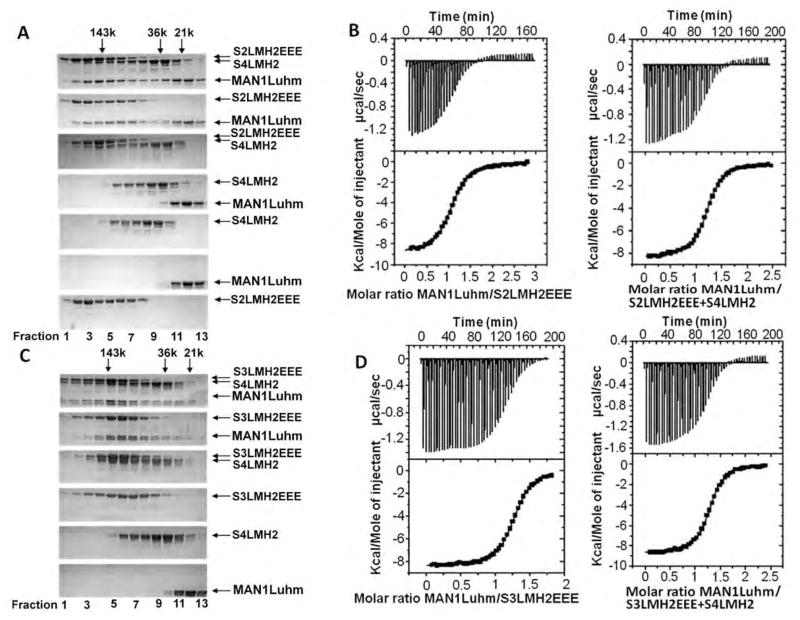
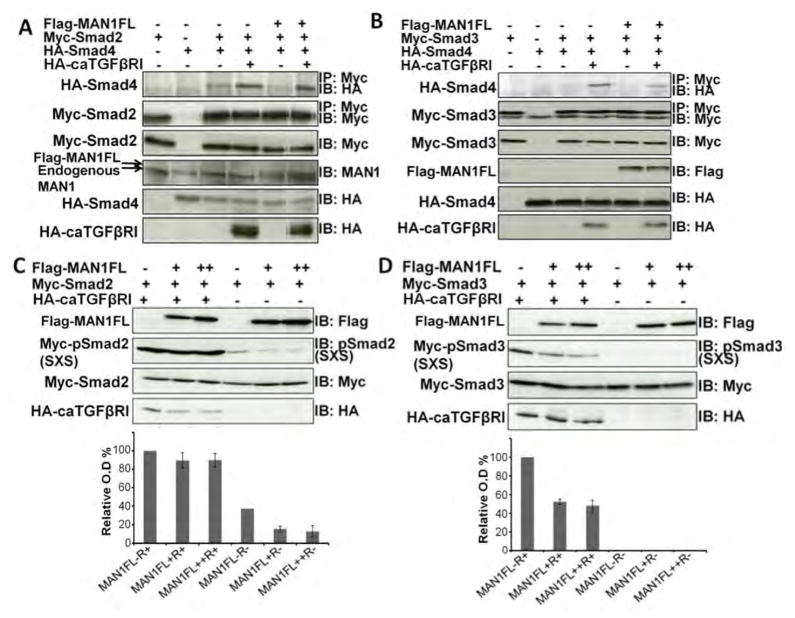
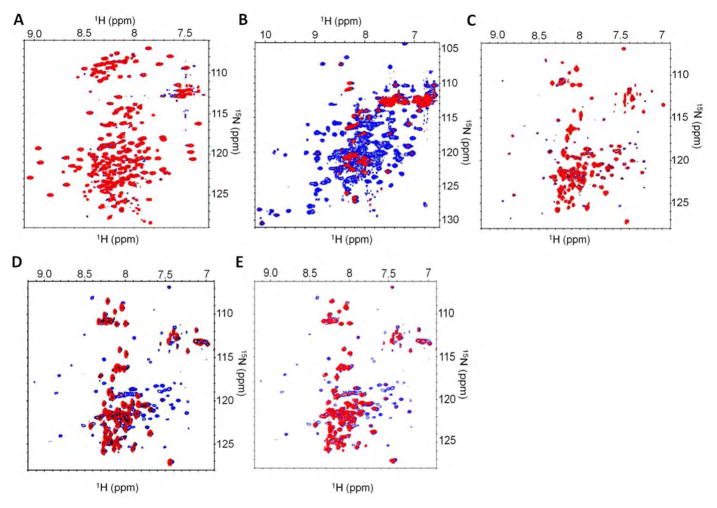
Signaling by transforming growth factor-β (TGF-β) is critical for various developmental processes and culminates in the activation of the transcription factors Smad2 and Smad3. MAN1, an integral protein of the inner nuclear membrane, inhibits TGF-β signaling by binding to Smad2 and Smad3. Depletion of the gene LEMD3 encoding MAN1 leads to developmental anomalies in mice, and heterozygous loss-of-function mutations in LEMD3 in humans cause sclerosing bone dysplasia. We modeled the three-dimensional structure of the MAN1-Smad2 complex from nuclear magnetic resonance and small-angle x-ray scattering data. As predicted by this model, we found that MAN1 competed in vitro and in cells with the transcription factor FAST1 (forkhead activin signal transducer 1) for binding to Smad2. The model further predicted that MAN1 bound to activated Smad2-Smad4 or Smad3-Smad4 complexes, which was confirmed by in vitro experiments; however, in cells, MAN1 bound only to Smad2 and Smad3 and not to the Smad4-containing complexes. Overexpression of MAN1 led to dephosphorylation of Smad2 and Smad3, thus hindering their recognition by Smad4, and MAN1 bound directly in vitro to the phosphatase PPM1A, which catalyzes the dephosphorylation of Smad2/3. These results demonstrate a nuclear envelope-localized mechanism of inactivating TGF-β signaling in which MAN1 competes with transcription factors for binding to Smad2 and Smad3 and facilitates their dephosphorylation by PPM1A.
Figures
Similar articles
-
MAN1, an integral protein of the inner nuclear membrane, binds Smad2 and Smad3 and antagonizes transforming growth factor-beta signaling.Hum Mol Genet. 2005 Feb 1;14(3):437-45. doi: 10.1093/hmg/ddi040. Epub 2004 Dec 15. Hum Mol Genet. 2005. PMID: 15601644
-
PPM1A dephosphorylates RanBP3 to enable efficient nuclear export of Smad2 and Smad3.EMBO Rep. 2011 Oct 28;12(11):1175-81. doi: 10.1038/embor.2011.174. EMBO Rep. 2011. PMID: 21960005 Free PMC article.
-
Structural analysis of the Smad2-MAN1 interaction that regulates transforming growth factor-β signaling at the inner nuclear membrane.Biochemistry. 2010 Sep 21;49(37):8020-32. doi: 10.1021/bi101153w. Biochemistry. 2010. PMID: 20715792
-
A phosphatase controls the fate of receptor-regulated Smads.Cell. 2006 Jun 2;125(5):838-40. doi: 10.1016/j.cell.2006.05.015. Cell. 2006. PMID: 16751094 Review.
-
A tale of two proteins: differential roles and regulation of Smad2 and Smad3 in TGF-beta signaling.J Cell Biochem. 2007 May 1;101(1):9-33. doi: 10.1002/jcb.21255. J Cell Biochem. 2007. PMID: 17340614 Review.
Cited by
-
Modulation of TGFbeta 2 levels by lamin A in U2-OS osteoblast-like cells: understanding the osteolytic process triggered by altered lamins.Oncotarget. 2015 Apr 10;6(10):7424-37. doi: 10.18632/oncotarget.3232. Oncotarget. 2015. PMID: 25823658 Free PMC article.
-
Diversity of Nuclear Lamin A/C Action as a Key to Tissue-Specific Regulation of Cellular Identity in Health and Disease.Front Cell Dev Biol. 2021 Oct 13;9:761469. doi: 10.3389/fcell.2021.761469. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34722546 Free PMC article. Review.
-
Molecular Pathology of Laminopathies.Annu Rev Pathol. 2022 Jan 24;17:159-180. doi: 10.1146/annurev-pathol-042220-034240. Epub 2021 Oct 21. Annu Rev Pathol. 2022. PMID: 34672689 Free PMC article. Review.
-
Genome-wide scan reveals LEMD3 and WIF1 on SSC5 as the candidates for porcine ear size.PLoS One. 2014 Jul 9;9(7):e102085. doi: 10.1371/journal.pone.0102085. eCollection 2014. PLoS One. 2014. PMID: 25006967 Free PMC article.
-
Posttranslational Regulation of Smads.Cold Spring Harb Perspect Biol. 2016 Dec 1;8(12):a022087. doi: 10.1101/cshperspect.a022087. Cold Spring Harb Perspect Biol. 2016. PMID: 27908935 Free PMC article. Review.
References
-
- Dauer WT, Worman HJ. The nuclear envelope as a signaling node in development and disease. Dev Cell. 2009;17:626–38. - PubMed
-
- Lin F, Blake DL, Callebaut I, Skerjanc IS, Holmer L, McBurney MW, Paulin-Levasseur M, Worman HJ. MAN1, an inner nuclear membrane protein that shares the LEM domain with lamina-associated polypeptide 2 and emerin. J Biol Chem. 2000;275:4840–7. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
