

The relative contributions of CYP3A4 and CYP3A5 to the metabolism of vinorelbine
- PMID: 23780963
- PMCID: PMC3876810
- DOI: 10.1124/dmd.113.051094
The relative contributions of CYP3A4 and CYP3A5 to the metabolism of vinorelbine
Abstract
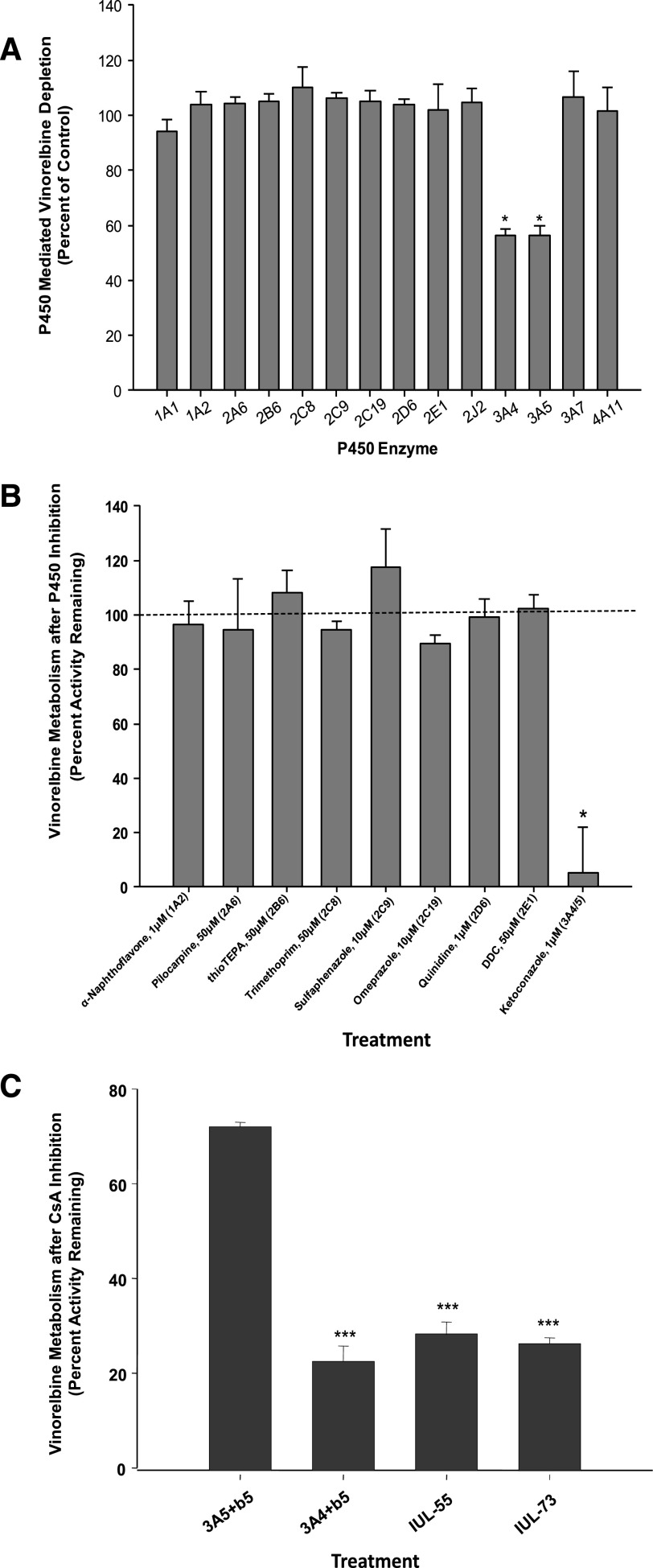
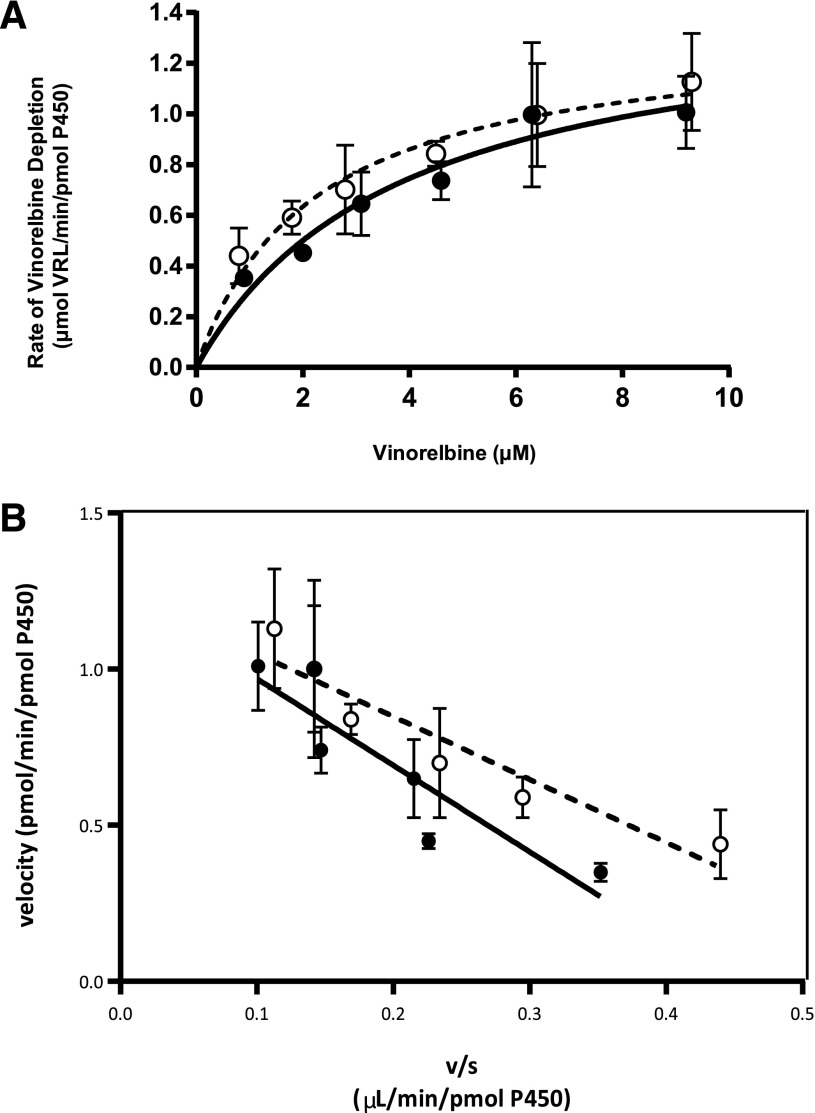
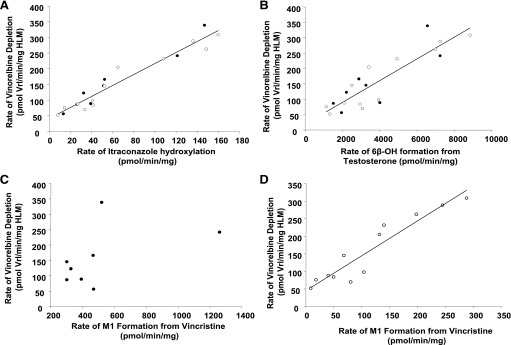
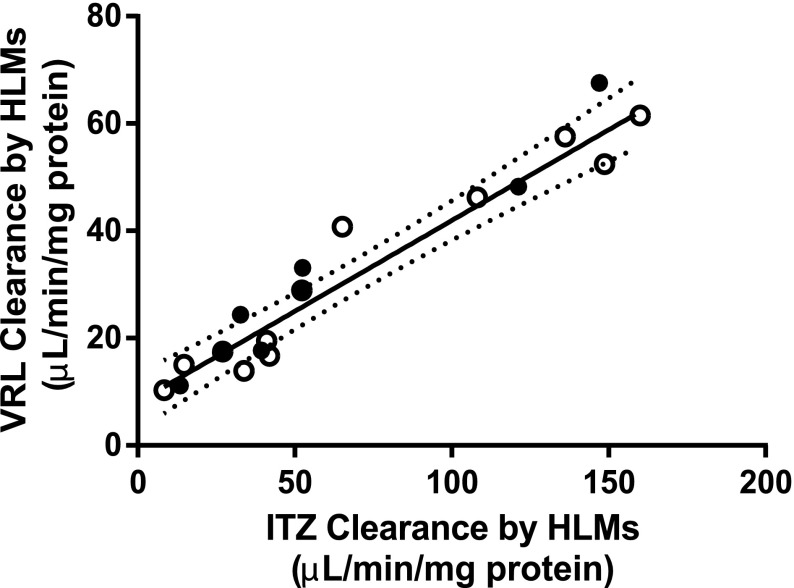
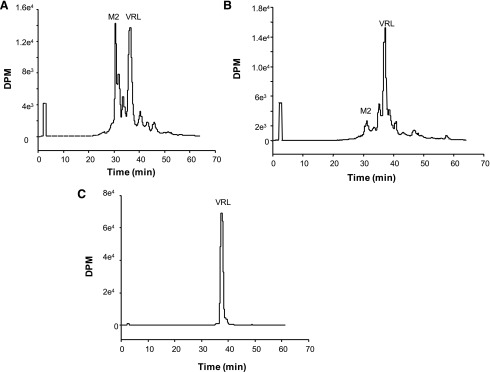
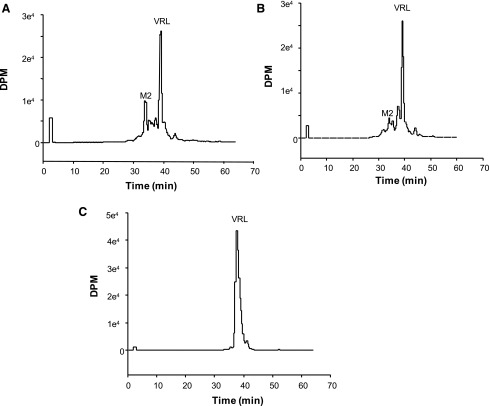
Vinorelbine is a semisynthetic vinca alkaloid used in the treatment of advanced breast and non-small cell lung cancers. Vincristine, a related vinca alkaloid, is 9-fold more efficiently metabolized by CYP3A5 than by CYP3A4 in vitro. This study quantified the relative contribution of CYP3A4 and CYP3A5 to the metabolism of vinorelbine in vitro using cDNA-expressed human cytochrome P450s (P450s) and human liver microsomes (HLMs). CYP3A4 and CYP3A5 were identified as the P450s capable of oxidizing vinorelbine using a panel of human enzymes and selective P450 inhibitors in HLMs. For CYP3A4 coexpressed with cytochrome b5 (CYP3A4+b5) and CYP3A5+b5, the Michaelis-Menten constants for vinorelbine were 2.6 and 3.6 μM, respectively, but the Vmax of 1.4 pmol/min/pmol was common to both enzymes. In HLMs, the intrinsic clearance of vinorelbine metabolism was highly correlated with CYP3A4 activity, and there was no significant difference in intrinsic clearance between CYP3A5 high and low expressers. When radiolabeled vinorelbine substrate was used, there were clear qualitative differences in metabolite formation fingerprints between CYP3A4+b5 and CYP3A5+b5 as determined by NMR and mass spectrometry analysis. One major metabolite (M2), a didehydro-vinorelbine, was present in both recombinant and microsomal systems but was more abundant in CYP3A4+b5 incubations. We conclude that despite the equivalent efficiency of recombinant CYP3A4 and CYP3A5 in vinorelbine metabolism the polymorphic expression of CYP3A5, as shown by the kinetics with HLMs, may have a minimal effect on systemic clearance of vinorelbine.
Figures
References
-
- Baker SD, Sparreboom A. (2006) Predicting vinorelbine disposition and toxicity: does BSA provide more than a “bad statistical association”? J Clin Oncol 24:2412–2413 - PubMed
-
- Bapiro TE, Egnell AC, Hasler JA, Masimirembwa CM. (2001) Application of higher throughput screening (HTS) inhibition assays to evaluate the interaction of antiparasitic drugs with cytochrome P450s. Drug Metab Dispos 29:30–35 - PubMed
-
- Beulz-Riché D, Grudé P, Puozzo C, Sautel F, Filaquier C, Riché C, Ratanasavanh D. (2005) Characterization of human cytochrome P450 isoenzymes involved in the metabolism of vinorelbine. Fundam Clin Pharmacol 19:545–553 - PubMed
-
- de Graeve J, van Heugen JC, Zorza G, Fahy J, Puozzo C. (2008) Metabolism pathway of vinorelbine (Navelbine) in human: characterisation of the metabolites by HPLC-MS/MS. J Pharm Biomed Anal 47:47–58 - PubMed
-
- Dennison JB, Jones DR, Renbarger JL, Hall SD. (2007) Effect of CYP3A5 expression on vincristine metabolism with human liver microsomes. J Pharmacol Exp Ther 321:553–563 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
