

The interaction between strigolactones and other plant hormones in the regulation of plant development
- PMID: 23785379
- PMCID: PMC3683633
- DOI: 10.3389/fpls.2013.00199
The interaction between strigolactones and other plant hormones in the regulation of plant development
Abstract
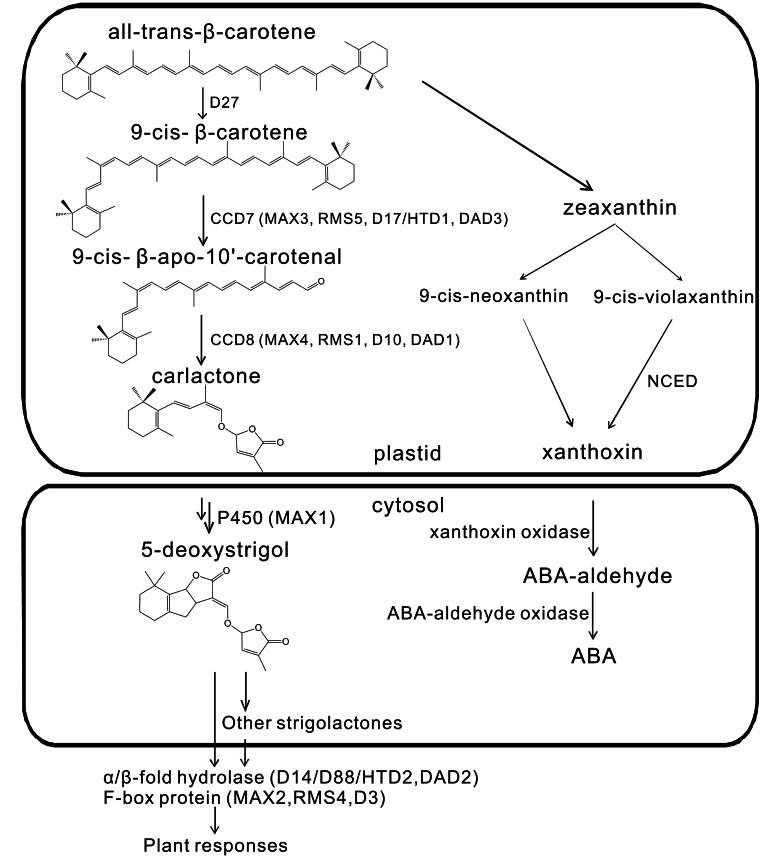
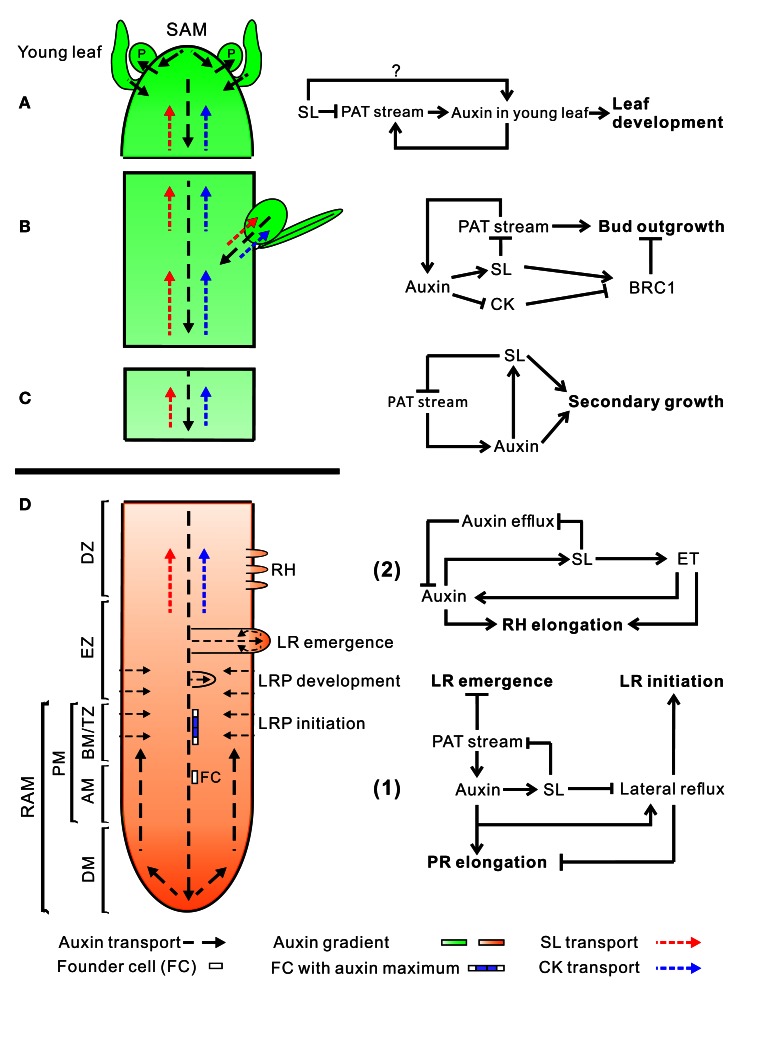
Plant hormones are small molecules derived from various metabolic pathways and are important regulators of plant development. The most recently discovered phytohormone class comprises the carotenoid-derived strigolactones (SLs). For a long time these compounds were only known to be secreted into the rhizosphere where they act as signaling compounds, but now we know they are also active as endogenous plant hormones and they have been in the spotlight ever since. The initial discovery that SLs are involved in the inhibition of axillary bud outgrowth, initiated a multitude of other studies showing that SLs also play a role in defining root architecture, secondary growth, hypocotyl elongation, and seed germination, mostly in interaction with other hormones. Their coordinated action enables the plant to respond in an appropriate manner to environmental factors such as temperature, shading, day length, and nutrient availability. Here, we will review the current knowledge on the crosstalk between SLs and other plant hormones-such as auxin, cytokinin, abscisic acid (ABA), ethylene (ET), and gibberellins (GA)-during different physiological processes. We will furthermore take a bird's eye view of how this hormonal crosstalk enables plants to respond to their ever changing environments.
Keywords: auxin; cytokinin; ethylene; gibberellins; hormone crosstalk; phenotypic plasticity; root and shoot architecture; strigolactone.
Figures
References
LinkOut - more resources
Full Text Sources
Other Literature Sources