

An excess-calcium-binding-site model predicts neurotransmitter release at the neuromuscular junction
- PMID: 23790384
- PMCID: PMC3686347
- DOI: 10.1016/j.bpj.2013.05.023
An excess-calcium-binding-site model predicts neurotransmitter release at the neuromuscular junction
Abstract
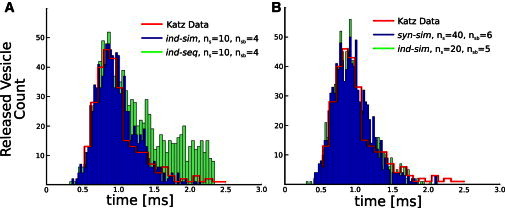
Despite decades of intense experimental studies, we still lack a detailed understanding of synaptic function. Fortunately, using computational approaches, we can obtain important new insights into the inner workings of these important neural systems. Here, we report the development of a spatially realistic computational model of an entire frog active zone in which we constrained model parameters with experimental data, and then used Monte Carlo simulation methods to predict the Ca(2+)-binding stoichiometry and dynamics that underlie neurotransmitter release. Our model reveals that 20-40 independent Ca(2+)-binding sites on synaptic vesicles, only a fraction of which need to bind Ca(2+) to trigger fusion, are sufficient to predict physiological release. Our excess-Ca(2+)-binding-site model has many functional advantages, agrees with recent data on synaptotagmin copy number, and is the first (to our knowledge) to link detailed physiological observations with the molecular machinery of Ca(2+)-triggered exocytosis. In addition, our model provides detailed microscopic insight into the underlying Ca(2+) dynamics during synapse activation.
Copyright © 2013 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
