

PI(4,5)P(2)-dependent and Ca(2+)-regulated ER-PM interactions mediated by the extended synaptotagmins
- PMID: 23791178
- PMCID: PMC3716012
- DOI: 10.1016/j.cell.2013.05.026
PI(4,5)P(2)-dependent and Ca(2+)-regulated ER-PM interactions mediated by the extended synaptotagmins
Abstract
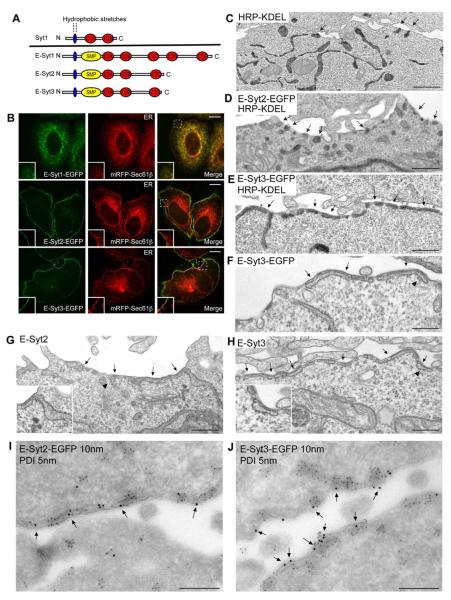
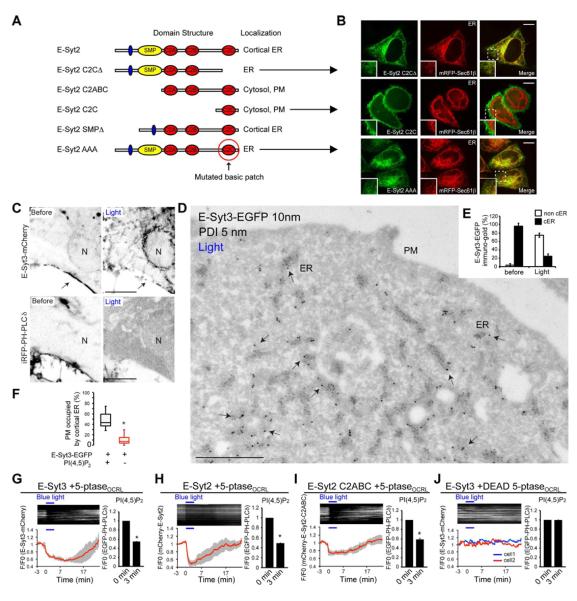
Most available information on endoplasmic reticulum (ER)-plasma membrane (PM) contacts in cells of higher eukaryotes concerns proteins implicated in the regulation of Ca(2+) entry. However, growing evidence suggests that such contacts play more general roles in cell physiology, pointing to the existence of additionally ubiquitously expressed ER-PM tethers. Here, we show that the three extended synaptotagmins (E-Syts) are ER proteins that participate in such tethering function via C2 domain-dependent interactions with the PM that require PI(4,5)P2 in the case of E-Syt2 and E-Syt3 and also elevation of cytosolic Ca(2+) in the case of E-Syt1. As they form heteromeric complexes, the E-Syts confer cytosolic Ca(2+) regulation to ER-PM contact formation. E-Syts-dependent contacts, however, are not required for store-operated Ca(2+) entry. Thus, the ER-PM tethering function of the E-Syts (tricalbins in yeast) mediates the formation of ER-PM contacts sites, which are functionally distinct from those mediated by STIM1 and Orai1.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures





Comment in
-
Inside-out connections: the ER meets the plasma membrane.Cell. 2013 Jun 20;153(7):1423-4. doi: 10.1016/j.cell.2013.05.054. Cell. 2013. PMID: 23791170 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
