

Role for ferredoxin:NAD(P)H oxidoreductase (FprA) in sulfate assimilation and siderophore biosynthesis in Pseudomonads
- PMID: 23794620
- PMCID: PMC3754593
- DOI: 10.1128/JB.00528-13
Role for ferredoxin:NAD(P)H oxidoreductase (FprA) in sulfate assimilation and siderophore biosynthesis in Pseudomonads
Abstract
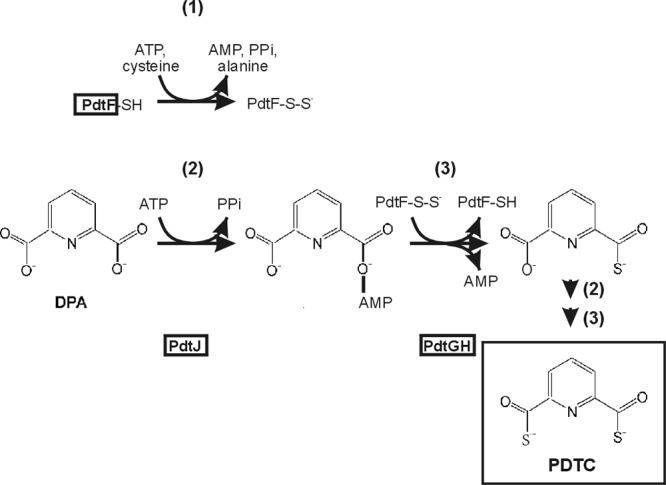
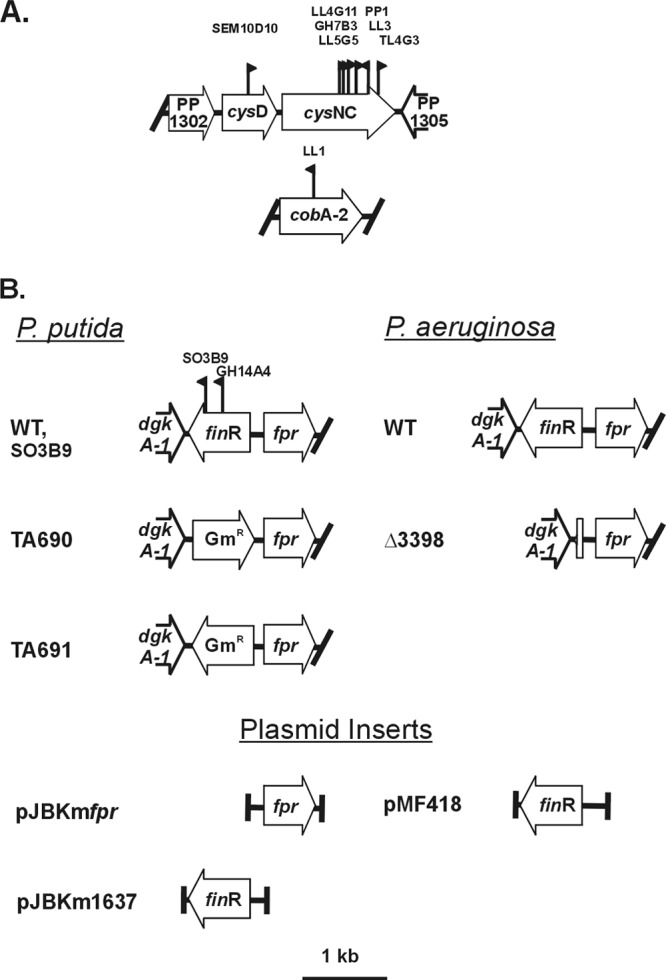
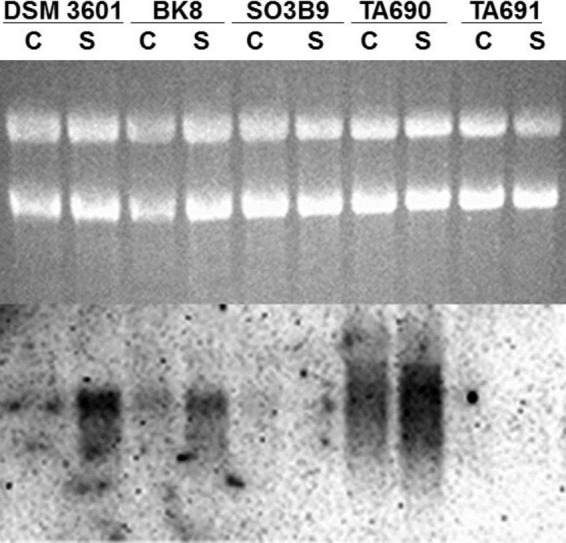
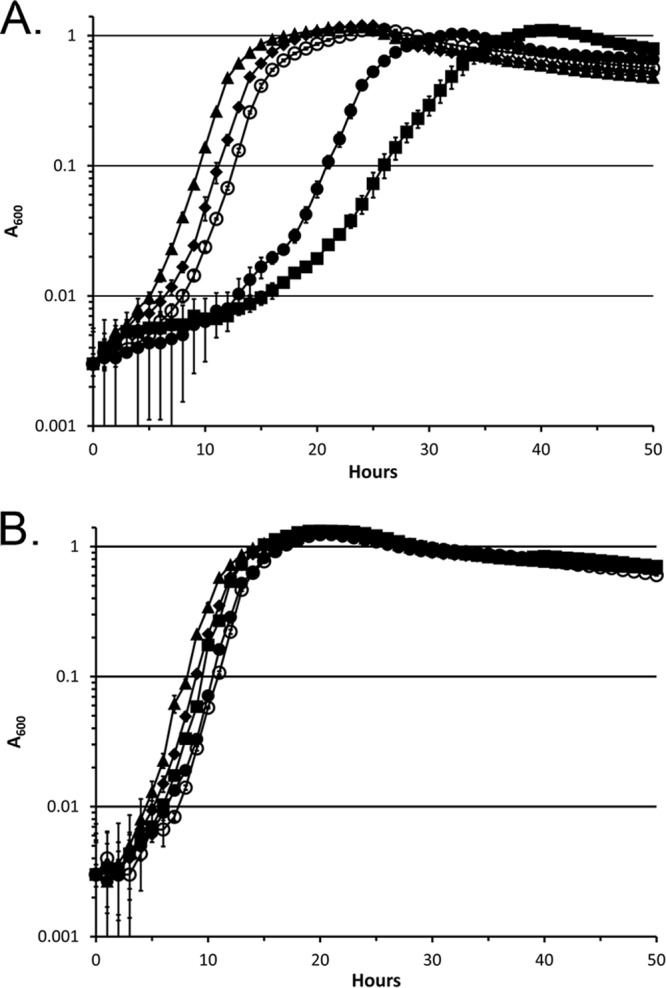
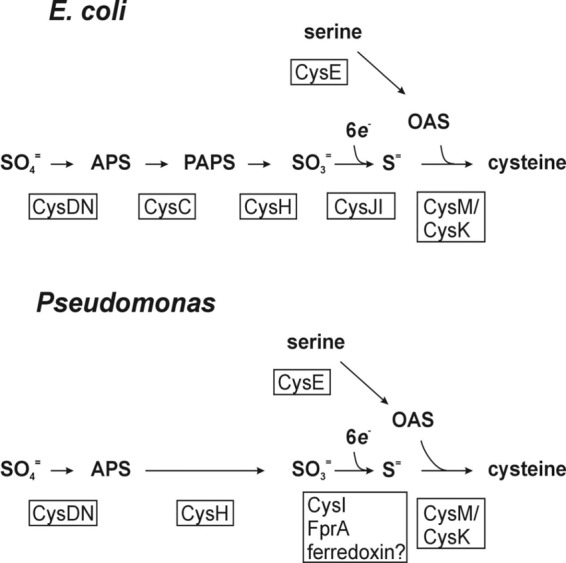
Pyridine-2,6-bis(thiocarboxylate) (PDTC), produced by certain pseudomonads, is a sulfur-containing siderophore that binds iron, as well as a wide range of transition metals, and it affects the net hydrolysis of the environmental contaminant carbon tetrachloride. The pathway of PDTC biosynthesis has not been defined. Here, we performed a transposon screen of Pseudomonas putida DSM 3601 to identify genes necessary for PDTC production (Pdt phenotype). Transposon insertions within genes for sulfate assimilation (cysD, cysNC, and cysG [cobA2]) dominated the collection of Pdt mutations. In addition, two insertions were within the gene for the LysR-type transcriptional activator FinR (PP1637). Phenotypic characterization indicated that finR mutants were cysteine bradytrophs. The Pdt phenotype of finR mutants could be complemented by the known target of FinR regulation, fprA (encoding ferredoxin:NADP(+) oxidoreductase), or by Escherichia coli cysJI (encoding sulfite reductase). These data indicate that fprA is necessary for effective sulfate assimilation by P. putida and that the effect of finR mutation on PDTC production was due to deficient expression of fprA and sulfite reduction. fprA expression in both P. putida and P. aeruginosa was found to be regulated by FinR, but in a manner dependent upon reduced sulfur sources, implicating FinR in sulfur regulatory physiology. The genes and phenotypes identified in this study indicated a strong dependence upon intracellular reduced sulfur/cysteine for PDTC biosynthesis and that pseudomonads utilize sulfite reduction enzymology distinct from that of E. coli and possibly similar to that of chloroplasts and other proteobacteria.
Figures
Similar articles
-
The FinR-regulated essential gene fprA, encoding ferredoxin NADP+ reductase: Roles in superoxide-mediated stress protection and virulence of Pseudomonas aeruginosa.PLoS One. 2017 Feb 10;12(2):e0172071. doi: 10.1371/journal.pone.0172071. eCollection 2017. PLoS One. 2017. PMID: 28187184 Free PMC article.
-
Molecular characterization of FinR, a novel redox-sensing transcriptional regulator in Pseudomonas putida KT2440.Microbiology (Reading). 2010 May;156(Pt 5):1487-1496. doi: 10.1099/mic.0.034181-0. Epub 2010 Jan 7. Microbiology (Reading). 2010. PMID: 20056701
-
Physiological and molecular genetic evaluation of the dechlorination agent, pyridine-2,6-bis(monothiocarboxylic acid) (PDTC) as a secondary siderophore of Pseudomonas.Environ Microbiol. 2004 Feb;6(2):159-69. doi: 10.1046/j.1462-2920.2003.00558.x. Environ Microbiol. 2004. PMID: 14756880
-
Identification and characterization of Pseudomonas membrane transporters necessary for utilization of the siderophore pyridine-2,6-bis(thiocarboxylic acid) (PDTC).Microbiology (Reading). 2006 Oct;152(Pt 10):3157-3166. doi: 10.1099/mic.0.29116-0. Microbiology (Reading). 2006. PMID: 17005994
-
Gene regulation of siderophore-mediated iron acquisition in Pseudomonas: not only the Fur repressor.Mol Microbiol. 1995 Aug;17(4):603-10. doi: 10.1111/j.1365-2958.1995.mmi_17040603.x. Mol Microbiol. 1995. PMID: 8801415 Review.
Cited by
-
An improved Tn7-lux reporter for broad host range, chromosomally-integrated promoter fusions in Gram-negative bacteria.J Microbiol Methods. 2015 Nov;118:75-7. doi: 10.1016/j.mimet.2015.08.016. Epub 2015 Sep 1. J Microbiol Methods. 2015. PMID: 26341612 Free PMC article.
-
NADPH-generating systems in bacteria and archaea.Front Microbiol. 2015 Jul 29;6:742. doi: 10.3389/fmicb.2015.00742. eCollection 2015. Front Microbiol. 2015. PMID: 26284036 Free PMC article. Review.
-
Pseudomonas aeruginosa IscR-Regulated Ferredoxin NADP(+) Reductase Gene (fprB) Functions in Iron-Sulfur Cluster Biogenesis and Multiple Stress Response.PLoS One. 2015 Jul 31;10(7):e0134374. doi: 10.1371/journal.pone.0134374. eCollection 2015. PLoS One. 2015. PMID: 26230408 Free PMC article.
-
Genomics Insights into Pseudomonas sp. CG01: An Antarctic Cadmium-Resistant Strain Capable of Biosynthesizing CdS Nanoparticles Using Methionine as S-Source.Genes (Basel). 2021 Jan 27;12(2):187. doi: 10.3390/genes12020187. Genes (Basel). 2021. PMID: 33514061 Free PMC article.
-
A High-Throughput Method for Identifying Novel Genes That Influence Metabolic Pathways Reveals New Iron and Heme Regulation in Pseudomonas aeruginosa.mSystems. 2021 Feb 2;6(1):e00933-20. doi: 10.1128/mSystems.00933-20. mSystems. 2021. PMID: 33531406 Free PMC article.
References
-
- Lee C-H, Lewis TA, Paszczynski A, Crawford RL. 1999. Identification of an extracellular catalyst of carbon tetrachloride dehalogenation from Pseudomonas stutzeri strain KC as pyridine-2,6-bis(thiocarboxylate). Biochem. Biophys. Res. Commun. 261:562–566 (Erratum, 265:770.) - PubMed
-
- Ockels W, Römer Budzikiewicz AH. 1978. An Fe(III) complex of pyridine-2,6-di-(monothiocarboxylic acid): a novel bacterial metabolic product. Tetrahedron Lett. 36:3341–3342
-
- Cortese M, Paszczynski A, Lewis TA, Sebat J, Crawford RL. 2002. Metal chelating properties of pyridine-2,6-bis(thiocarboxylic acid) produced by Pseudomonas spp. and the biological activities of the formed complexes. Biometals 15:103–120 - PubMed
-
- Leach L, Lewis TA. 2007. The role of the siderophore pyridine-2,6-bis (thiocarboxylic acid) (PDTC) in zinc utilization by Pseudomonas putida DSM 3601. Biometals 20:717–726 - PubMed
-
- Lewis TA, Paszczynski A, Gordon-Wylie S, Jeedigunta S, Lee C-H, Crawford RL. 2001. Carbon tetrachloride dechlorination by the bacterial transition metal chelator pyridine-2,6-bis(thiocarboxylic acid). Environ. Sci. Technol. 35:552–559 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
