

The Arp2/3 complex mediates multigeneration dendritic protrusions for efficient 3-dimensional cancer cell migration
- PMID: 23796785
- PMCID: PMC4046187
- DOI: 10.1096/fj.12-224352
The Arp2/3 complex mediates multigeneration dendritic protrusions for efficient 3-dimensional cancer cell migration
Abstract
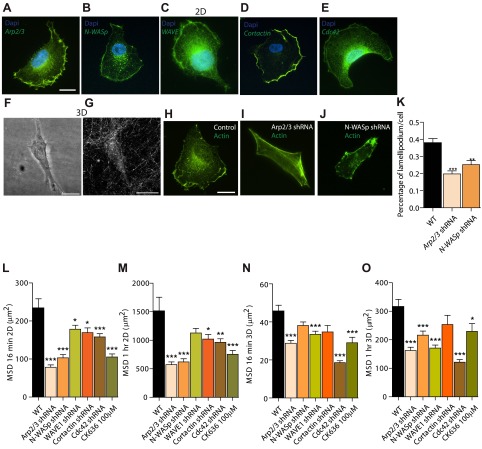
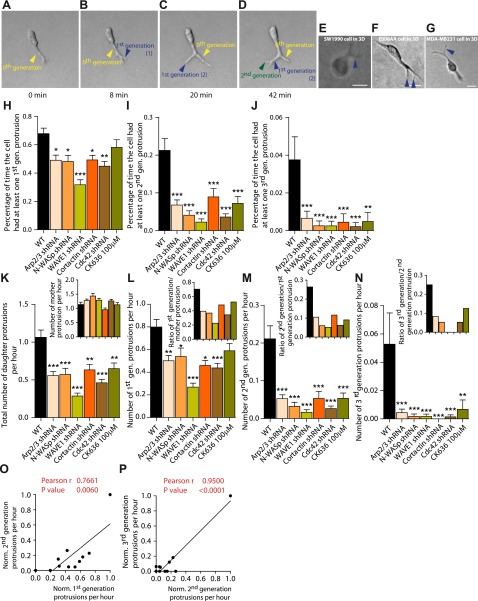
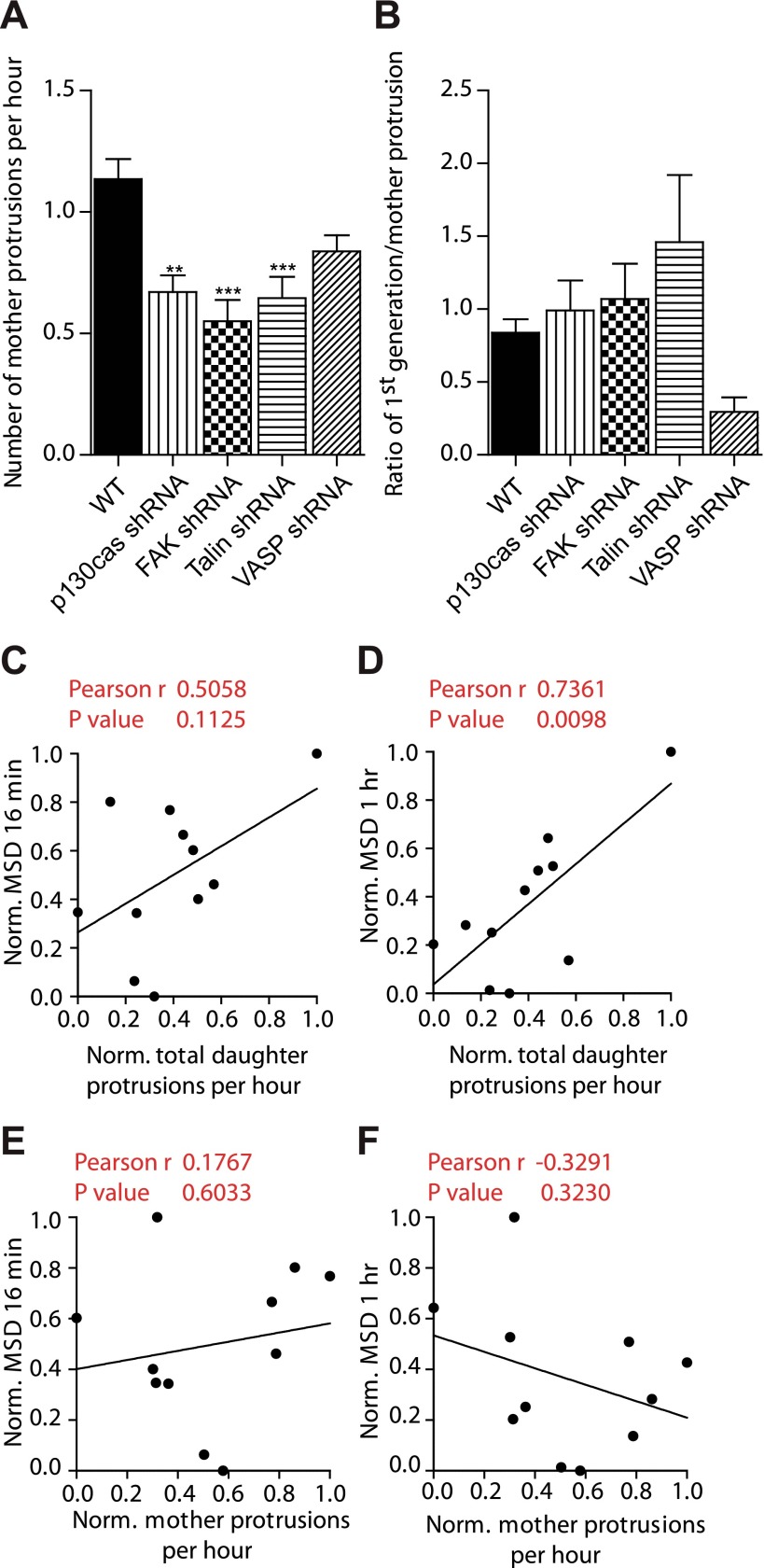
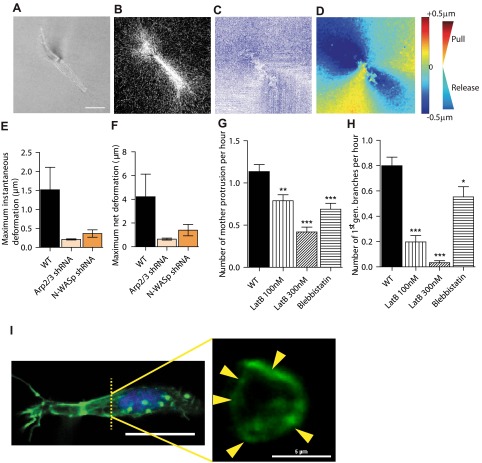
Arp2/3 is a protein complex that nucleates actin filament assembly in the lamellipodium in adherent cells crawling on planar 2-dimensional (2D) substrates. However, in physiopathological situations, cell migration typically occurs within a 3-dimensional (3D) environment, and little is known about the role of Arp2/3 and associated proteins in 3D cell migration. Using time resolved live-cell imaging and HT1080, a fibrosarcoma cell line commonly used to study cell migration, we find that the Arp2/3 complex and associated proteins N-WASP, WAVE1, cortactin, and Cdc42 regulate 3D cell migration. We report that this regulation is caused by formation of multigeneration dendritic protrusions, which mediate traction forces on the surrounding matrix and effective cell migration. The primary protrusions emanating directly from the cell body and prolonging the nucleus forms independent of Arp2/3 and dependent on focal adhesion proteins FAK, talin, and p130Cas. The Arp2/3 complex, N-WASP, WAVE1, cortactin, and Cdc42 regulate the secondary protrusions branching off from the primary protrusions. In 3D matrices, fibrosarcoma cells as well as migrating breast, pancreatic, and prostate cancer cells do not display lamellipodial structures. This study characterizes the unique topology of protrusions made by cells in a 3D matrix and show that these dendritic protrusions play a critical role in 3D cell motility and matrix deformation. The relative contribution of these proteins to 3D migration is significantly different from their role in 2D migration.
Keywords: 3D environment; collagen I matrix; matrix deformation.
Figures
References
-
- Pollard T. D., Borisy G. G. (2003) Cellular motility driven by assembly and disassembly of actin filaments. Cell 112, 453–465 - PubMed
-
- Goley E. D., Welch M. D. (2006) The ARP2/3 complex: an actin nucleator comes of age. Nat. Rev. Mol. Cell Biol. 7, 713–726 - PubMed
-
- Derry J. M., Ochs H. D., Francke U. (1994) Isolation of a novel gene mutated in Wiskott-Aldrich syndrome. Cell 78, 635–644 - PubMed
-
- Kim A. S., Kakalis L. T., Abdul-Manan N., Liu G. A., Rosen M. K. (2000) Autoinhibition and activation mechanisms of the Wiskott-Aldrich syndrome protein. Nature 404, 151–158 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
