

An IKKα-E2F1-BMI1 cascade activated by infiltrating B cells controls prostate regeneration and tumor recurrence
- PMID: 23796898
- PMCID: PMC3713424
- DOI: 10.1101/gad.220202.113
An IKKα-E2F1-BMI1 cascade activated by infiltrating B cells controls prostate regeneration and tumor recurrence
Abstract
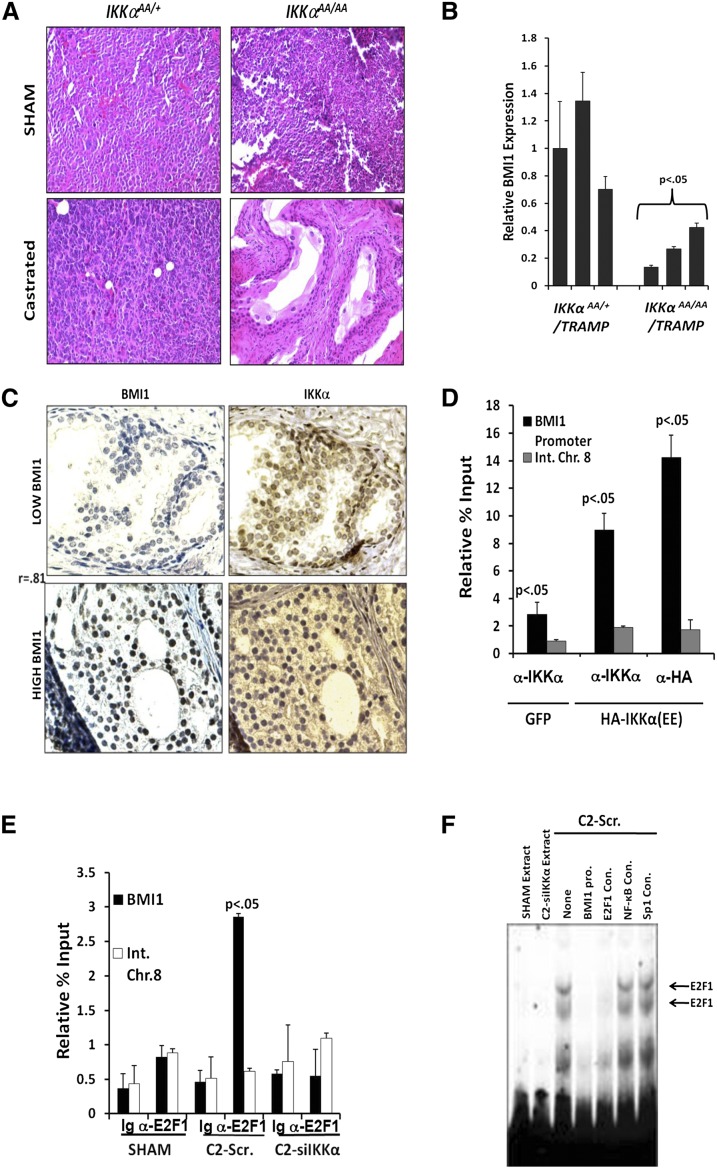
Androgen-deprived prostate cancer (PCa) is infiltrated by B lymphocytes that produce cytokines that activate IκB kinase α (IKKα) to accelerate the emergence of castration-resistant tumors. We now demonstrate that infiltrating B lymphocytes and IKKα are also required for androgen-dependent expansion of epithelial progenitors responsible for prostate regeneration. In these cells and in PCa cells, IKKα phosphorylates transcription factor E2F1 on a site that promotes its nuclear translocation, association with the coactivator CBP, and recruitment to critical genomic targets that include Bmi1, a key regulator of normal and cancerous prostate stem cell renewal. The IKKα-BMI1 pathway is also activated in human PCa.
Keywords: B cells; prostate; regeneration.
Figures




References
-
- Beachy PA, Karhadkar SS, Berman DM 2004. Tissue repair and stem cell renewal in carcinogenesis. Nature 432: 324–331 - PubMed
-
- Ben-Neriah Y, Karin M 2011. Inflammation meets cancer, with NF-κB as the matchmaker. Nat Immunol 12: 715–723 - PubMed
-
- Cao Y, Bonizzi G, Seagroves TN, Greten FR, Johnson R, Schmidt EV, Karin M 2001. IKKα provides an essential link between RANK signaling and cyclin D1 expression during mammary gland development. Cell 107: 763–775 - PubMed
-
- Chen J, Trounstine M, Alt FW, Young F, Kurahara C, Loring JF, Huszar D 1993. Immunoglobulin gene rearrangement in B cell deficient mice generated by targeted deletion of the JH locus. Int Immunol 5: 647–656 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials