

Identification of beryllium-dependent peptides recognized by CD4+ T cells in chronic beryllium disease
- PMID: 23797096
- PMCID: PMC3698527
- DOI: 10.1084/jem.20122426
Identification of beryllium-dependent peptides recognized by CD4+ T cells in chronic beryllium disease
Abstract
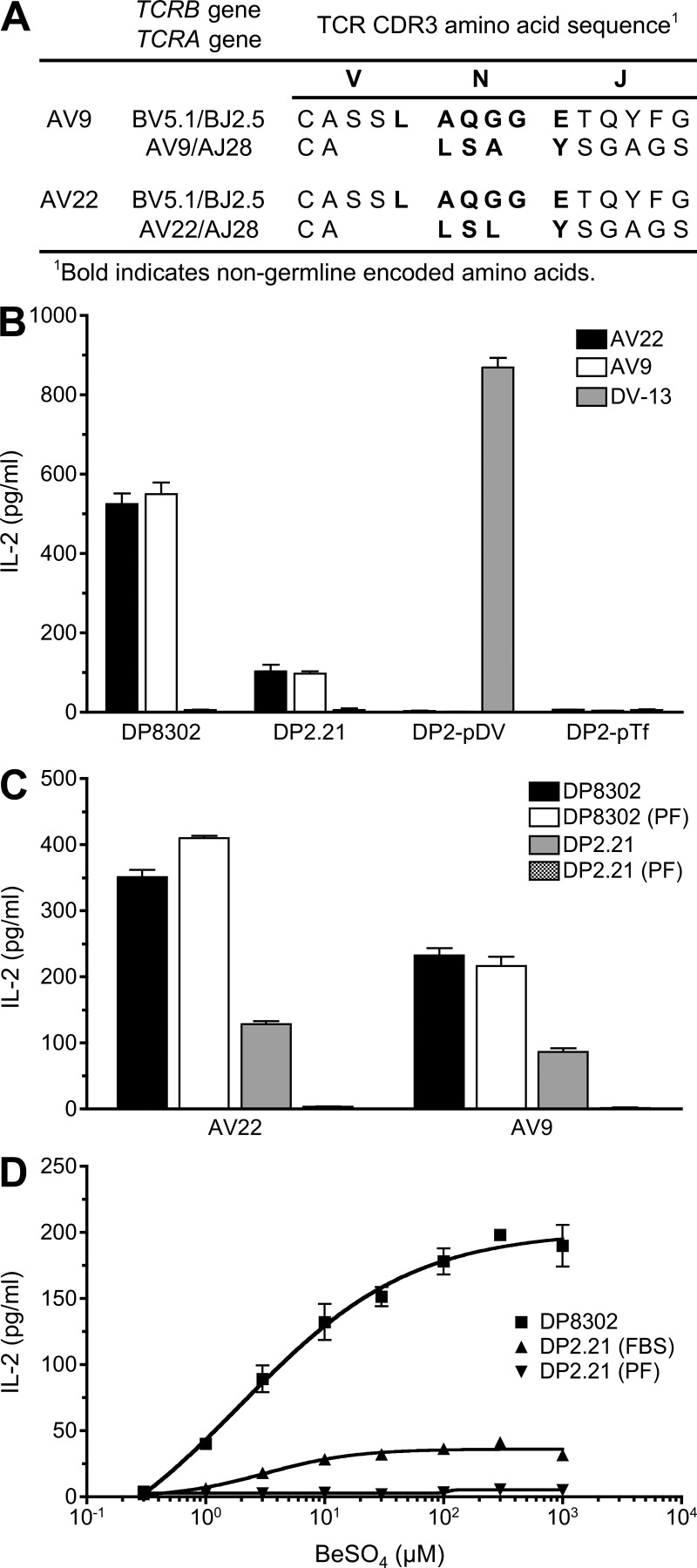
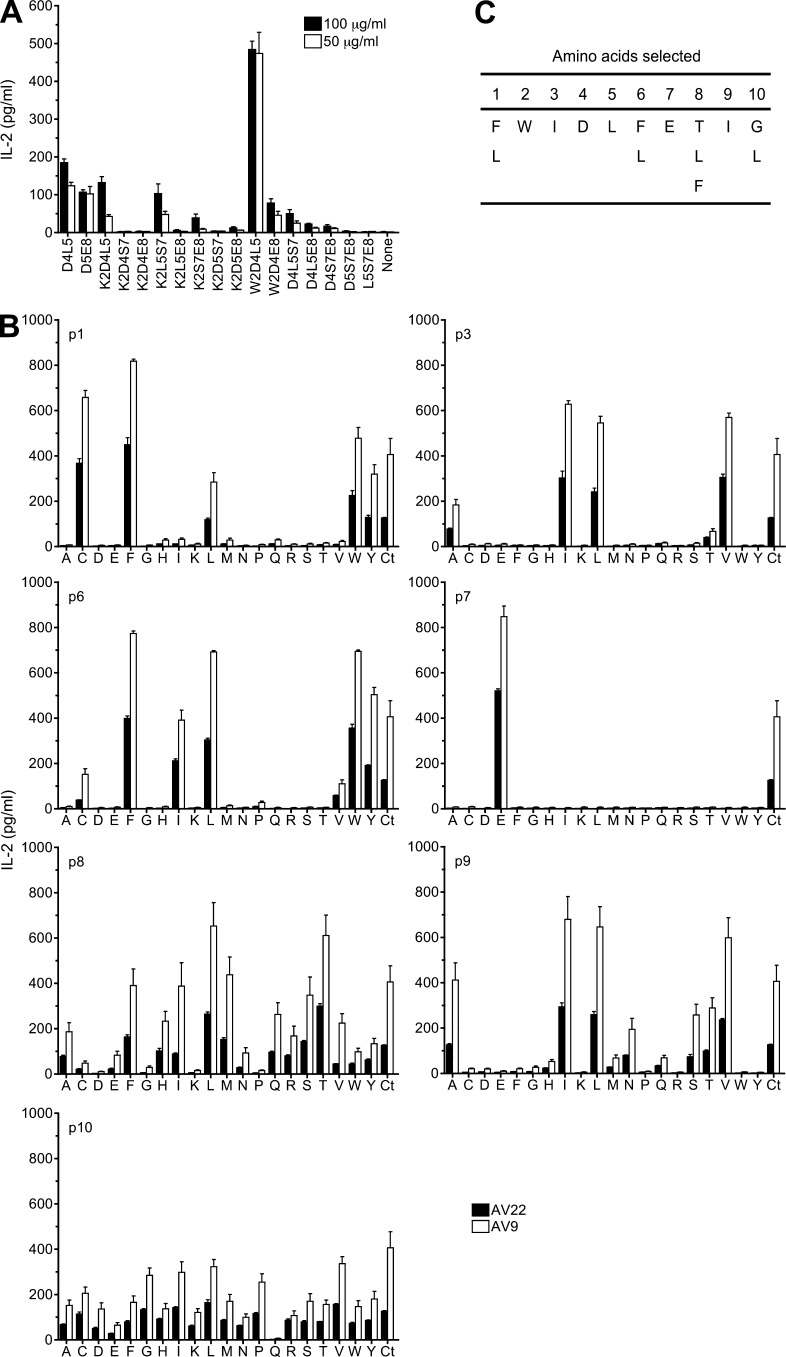
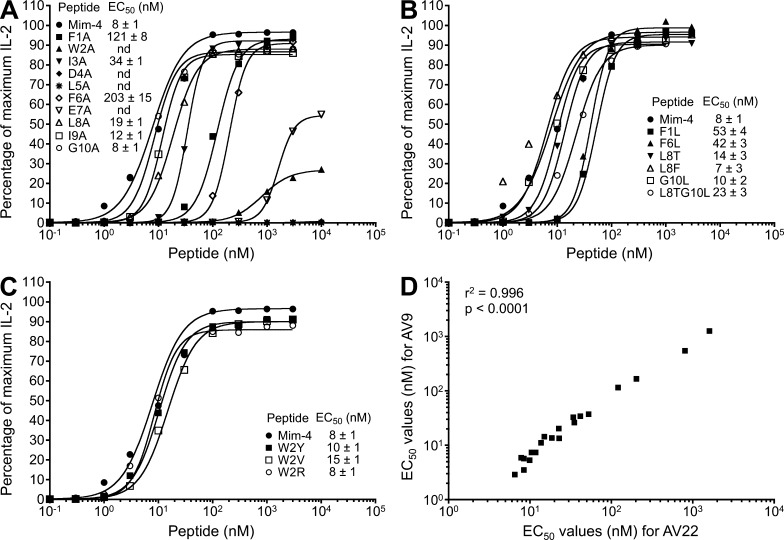
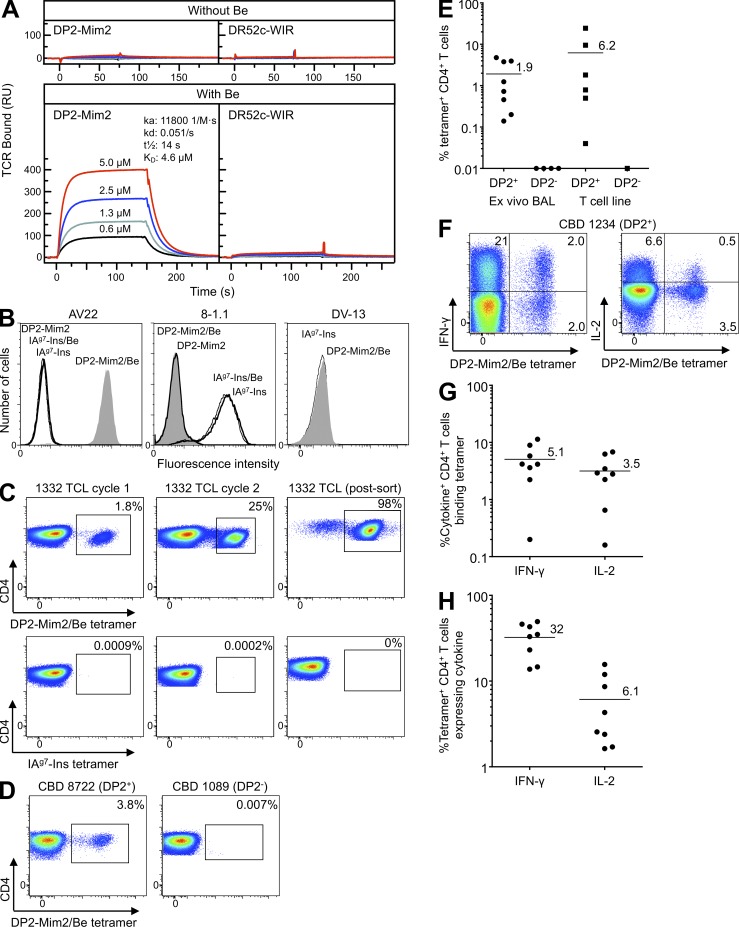
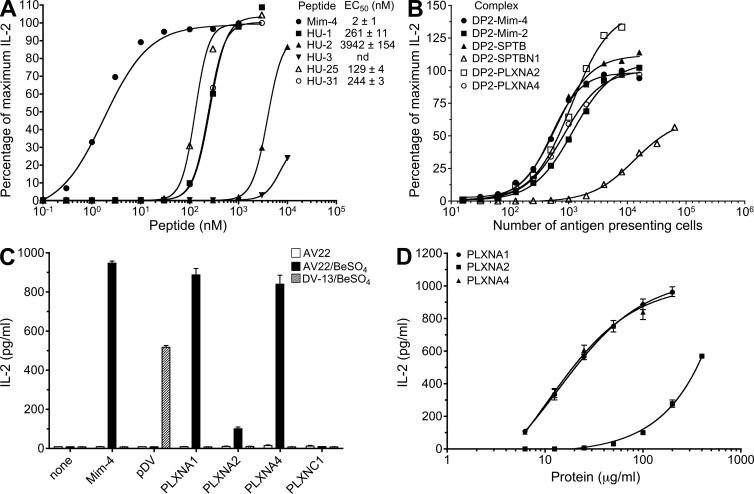
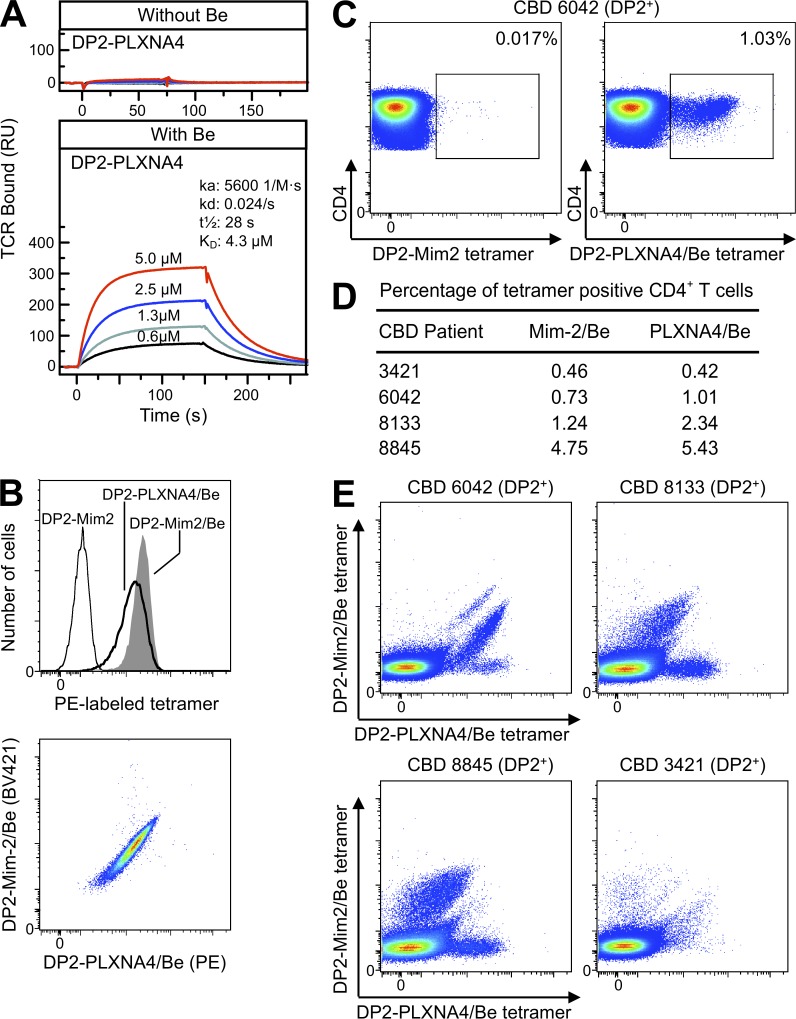
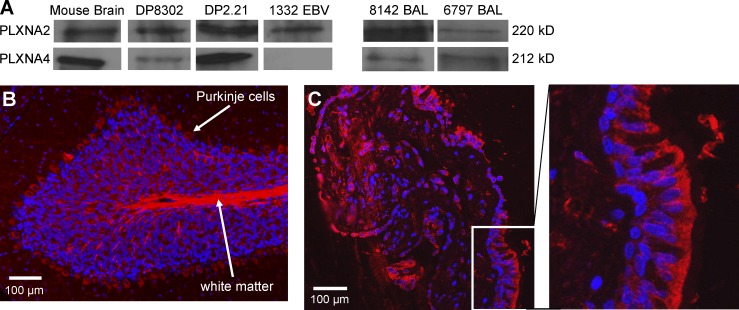
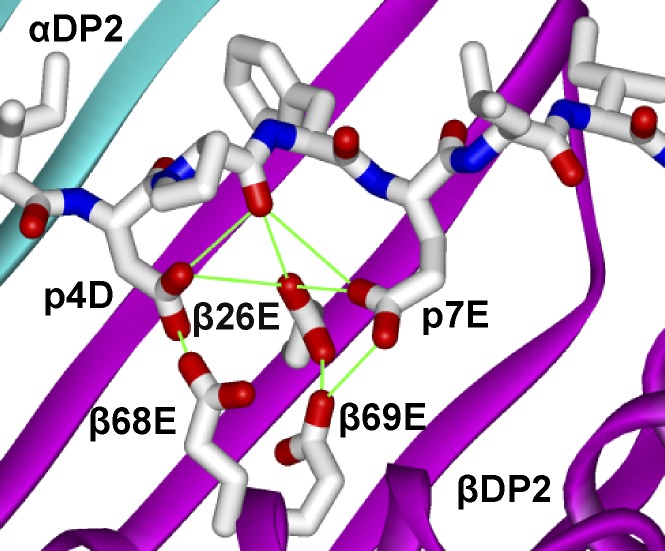
Chronic beryllium disease (CBD) is a granulomatous disorder characterized by an influx of beryllium (Be)-specific CD4⁺ T cells into the lung. The vast majority of these T cells recognize Be in an HLA-DP–restricted manner, and peptide is required for T cell recognition. However, the peptides that stimulate Be-specific T cells are unknown. Using positional scanning libraries and fibroblasts expressing HLA-DP2, the most prevalent HLA-DP molecule linked to disease, we identified mimotopes and endogenous self-peptides that bind to MHCII and Be, forming a complex recognized by pathogenic CD4⁺ T cells in CBD. These peptides possess aspartic and glutamic acid residues at p4 and p7, respectively, that surround the putative Be-binding site and cooperate with HLA-DP2 in Be coordination. Endogenous plexin A peptides and proteins, which share the core motif and are expressed in lung, also stimulate these TCRs. Be-loaded HLA-DP2–mimotope and HLA-DP2–plexin A4 tetramers detected high frequencies of CD4⁺ T cells specific for these ligands in all HLADP2+ CBD patients tested. Thus, our findings identify the first ligand for a CD4⁺ T cell involved in metal-induced hypersensitivity and suggest a unique role of these peptides in metal ion coordination and the generation of a common antigen specificity in CBD.
Figures
References
-
- Bill J.R., Mack D.G., Falta M.T., Maier L.A., Sullivan A.K., Joslin F.G., Martin A.K., Freed B.M., Kotzin B.L., Fontenot A.P. 2005. Beryllium presentation to CD4+ T cells is dependent on a single amino acid residue of the MHC class II β-chain. J. Immunol. 175:7029–7037 - PubMed
-
- Boen E., Crownover A.R., McIlhaney M., Korman A.J., Bill J. 2000. Identification of T cell ligands in a library of peptides covalently attached to HLA-DR4. J. Immunol. 165:2040–2047 - PubMed
-
- Crawford F., Stadinski B., Jin N., Michels A., Nakayama M., Pratt P., Marrack P., Eisenbarth G., Kappler J.W. 2011. Specificity and detection of insulin-reactive CD4+ T cells in type 1 diabetes in the nonobese diabetic (NOD) mouse. Proc. Natl. Acad. Sci. USA. 108:16729–16734 10.1073/pnas.1113954108 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
