

Structural basis of cargo recognitions for class V myosins
- PMID: 23798443
- PMCID: PMC3710815
- DOI: 10.1073/pnas.1306768110
Structural basis of cargo recognitions for class V myosins
Abstract
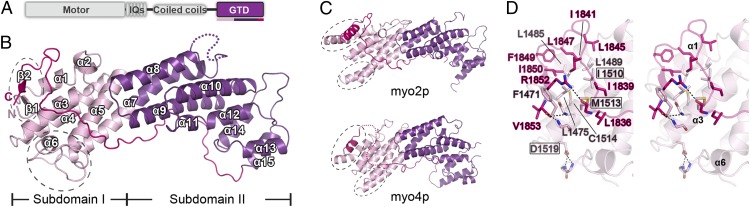
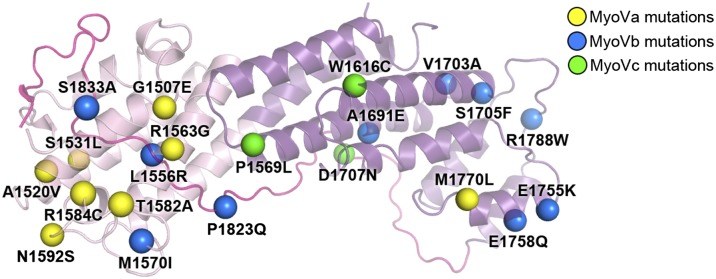
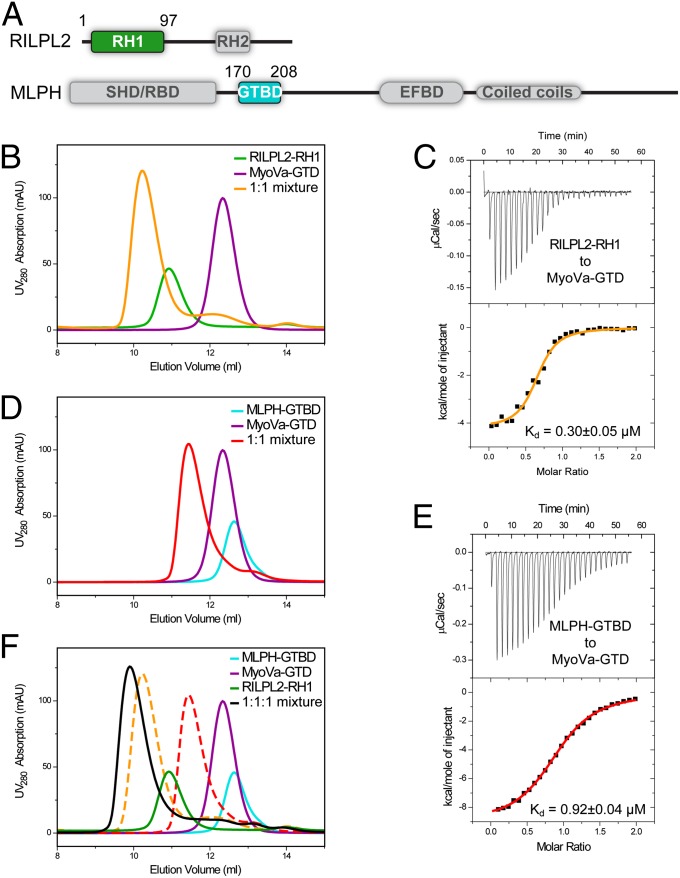
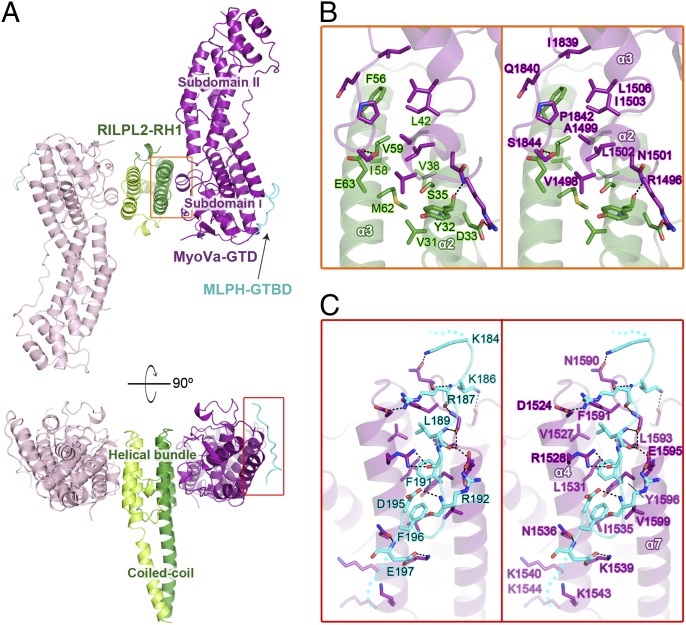
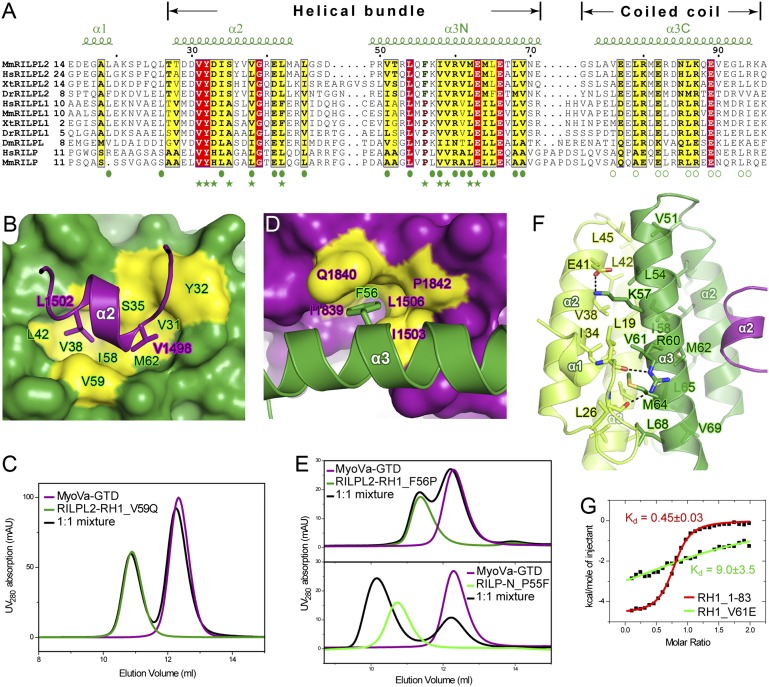
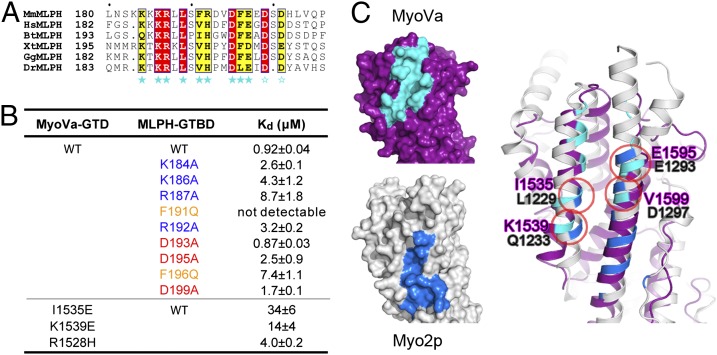
Class V myosins (MyoV), the most studied unconventional myosins, recognize numerous cargos mainly via the motor's globular tail domain (GTD). Little is known regarding how MyoV-GTD recognizes such a diverse array of cargos specifically. Here, we solved the crystal structures of MyoVa-GTD in its apo-form and in complex with two distinct cargos, melanophilin and Rab interacting lysosomal protein-like 2. The apo-MyoVa-GTD structure indicates that most mutations found in patients with Griscelli syndrome, microvillus inclusion disease, or cancers or in "dilute" rodents likely impair the folding of GTD. The MyoVa-GTD/cargo complex structure reveals two distinct cargo-binding surfaces, one primarily via charge-charge interaction and the other mainly via hydrophobic interactions. Structural and biochemical analysis reveal the specific cargo-binding specificities of various isoforms of mammalian MyoV as well as very different cargo recognition mechanisms of MyoV between yeast and higher eukaryotes. The MyoVa-GTD structures resolved here provide a framework for future functional studies of vertebrate class V myosins.
Keywords: MLPH; RILPL2; granuphilin; myo5a; myosin V.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Mercer JA, Seperack PK, Strobel MC, Copeland NG, Jenkins NA. Novel myosin heavy chain encoded by murine dilute coat colour locus. Nature. 1991;349(6311):709–713. - PubMed
-
- Hammer JA, 3rd, Sellers JR. Walking to work: Roles for class V myosins as cargo transporters. Nat Rev Mol Cell Biol. 2012;13(1):13–26. - PubMed
-
- Sellers JR, Weisman LS. Myosin V. In: Coluccio LM, editor. Myosins, a Superfamily of Molecular Motors. Vol 9. Dordrecht: Springer; 2008. pp. 289–323.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
