

Altered innate defenses in the neonatal gastrointestinal tract in response to colonization by neuropathogenic Escherichia coli
- PMID: 23798529
- PMCID: PMC3754193
- DOI: 10.1128/IAI.00268-13
Altered innate defenses in the neonatal gastrointestinal tract in response to colonization by neuropathogenic Escherichia coli
Abstract
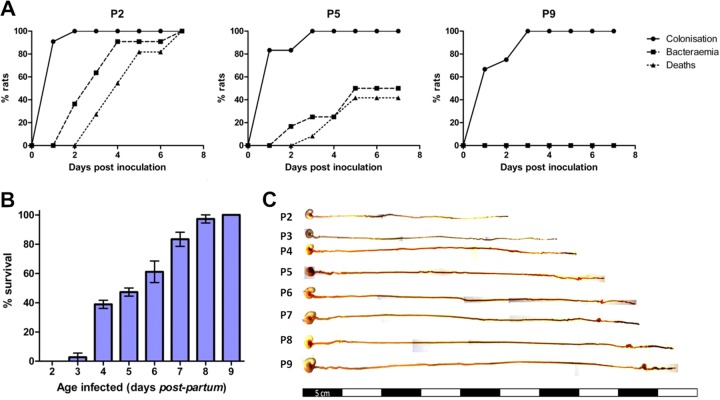
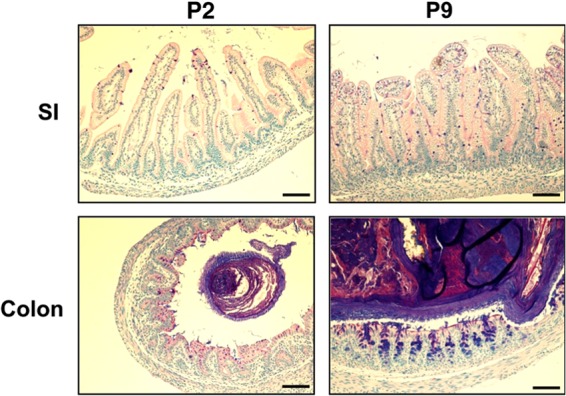
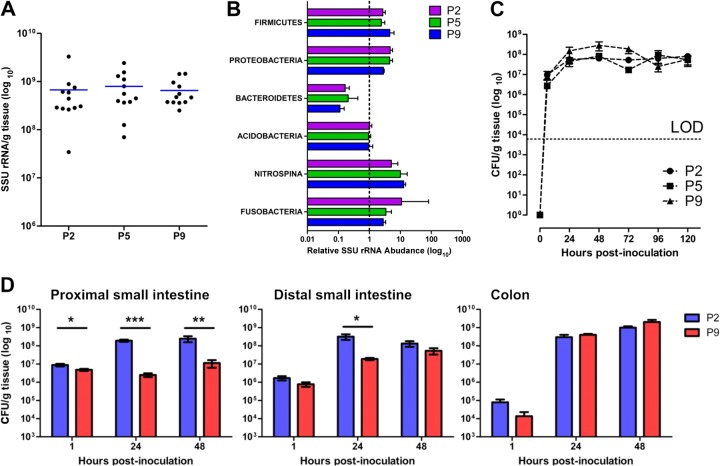
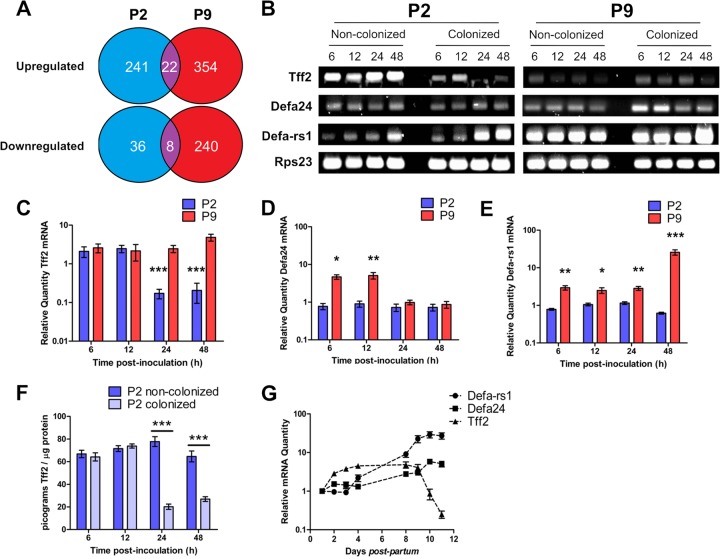
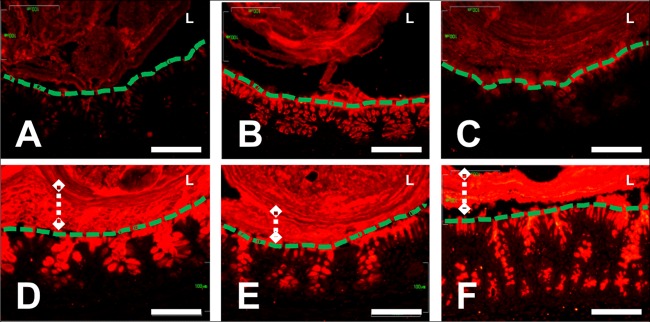
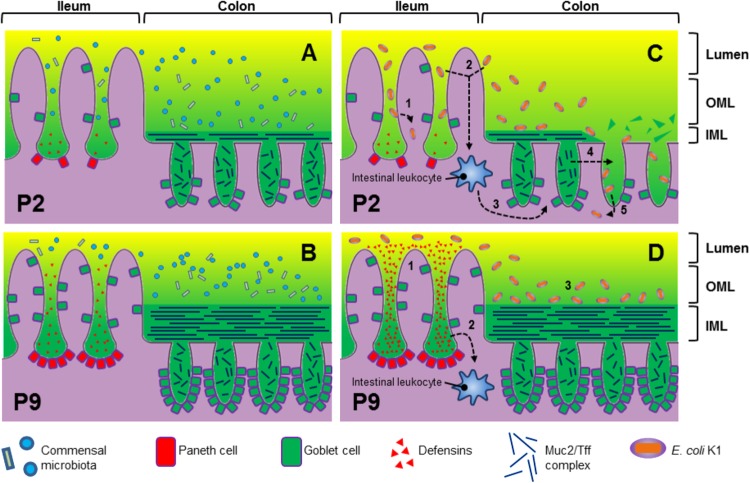
Two-day-old (P2), but not 9-day-old (P9), rat pups are susceptible to systemic infection following gastrointestinal colonization by Escherichia coli K1. Age dependency reflects the capacity of colonizing K1 to translocate from gastrointestinal (GI) tract to blood. A complex GI microbiota developed by P2, showed little variation over P2 to P9, and did not prevent stable K1 colonization. Substantial developmental expression was observed over P2 to P9, including upregulation of genes encoding components of the small intestinal (α-defensins Defa24 and Defa-rs1) and colonic (trefoil factor Tff2) mucus barrier. K1 colonization modulated expression of these peptides: developmental expression of Tff2 was dysregulated in P2 tissues and was accompanied by a decrease in mucin Muc2. Conversely, α-defensin genes were upregulated in P9 tissues. We propose that incomplete development of the mucus barrier during early neonatal life and the capacity of colonizing K1 to interfere with mucus barrier maturation provide opportunities for neuropathogen translocation into the bloodstream.
Figures
References
-
- Harvey D, Holt DE, Bedford H. 1999. Bacterial meningitis in the newborn: a prospective study of mortality and morbidity. Semin. Perinatol. 23:218–225 - PubMed
-
- Robbins JB, McCracken GH, Gotschlich EC, Ørskov F, Ørskov I, Hanson LA. 1974. Escherichia coli K1 polysaccharide associated with neonatal meningitis. N. Engl. J. Med. 290:1216–1220 - PubMed
-
- Rutishauser U. 2008. Polysialic acid in the plasticity of the developing and adult vertebrate nervous system. Nat. Rev. Neurosci. 9:26–35 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
