

Inhibition of HDAC6 deacetylase activity increases its binding with microtubules and suppresses microtubule dynamic instability in MCF-7 cells
- PMID: 23798680
- PMCID: PMC3829339
- DOI: 10.1074/jbc.M113.489328
Inhibition of HDAC6 deacetylase activity increases its binding with microtubules and suppresses microtubule dynamic instability in MCF-7 cells
Abstract
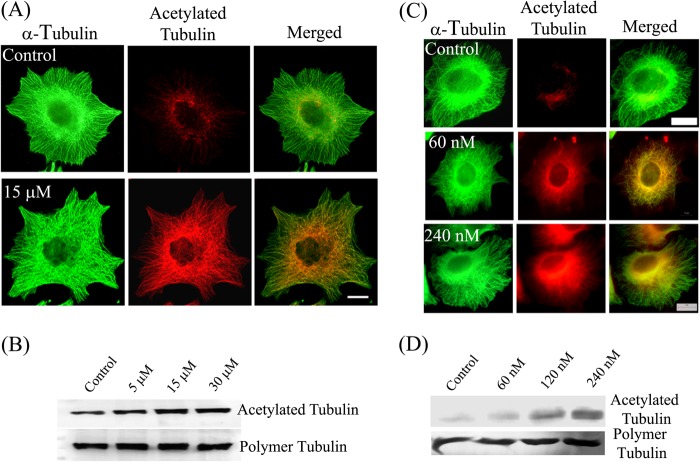
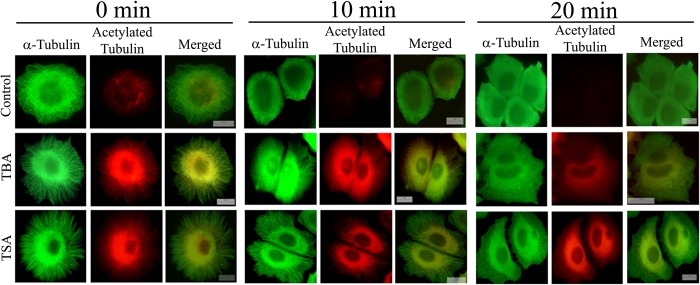
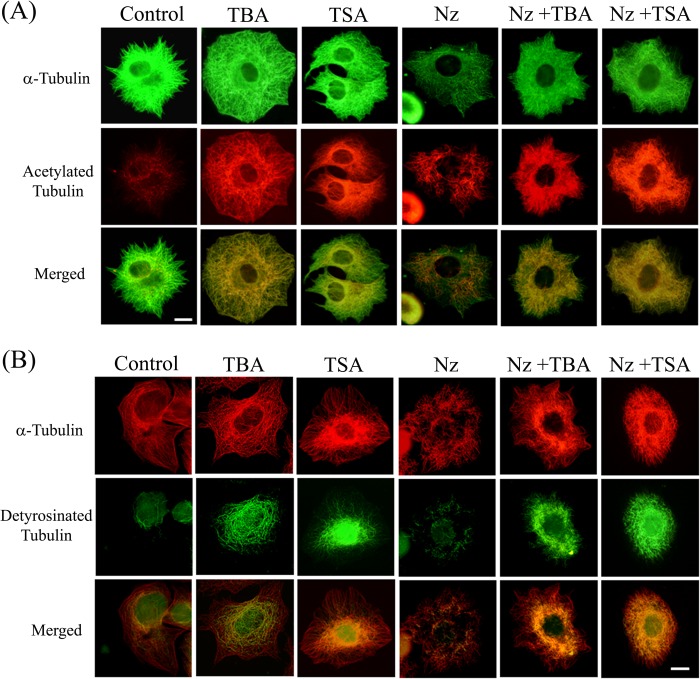
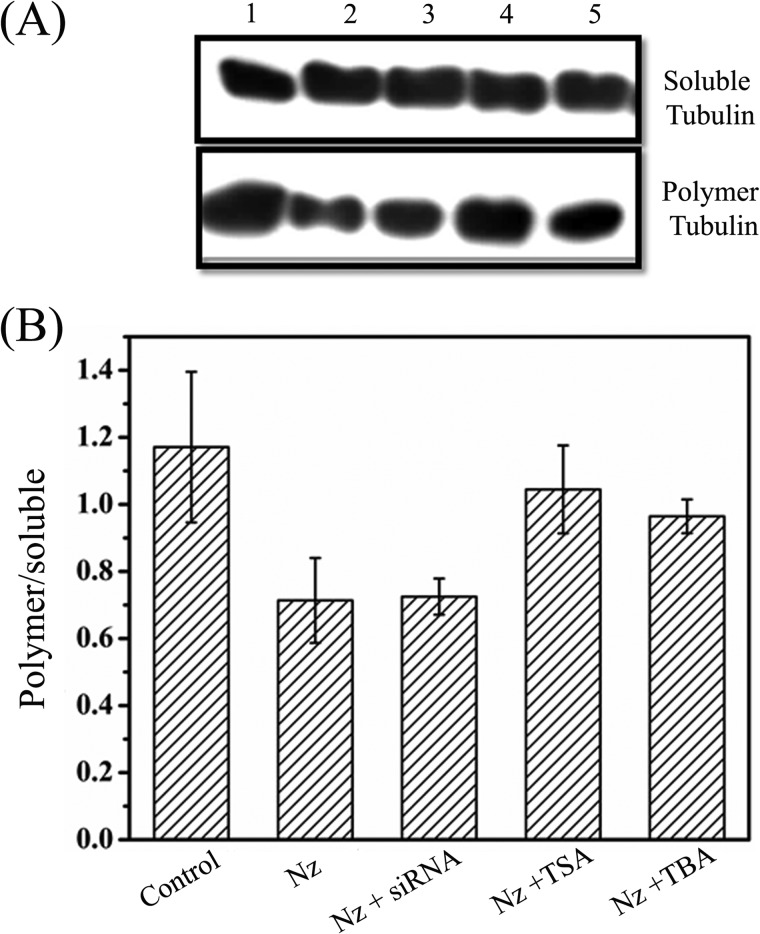
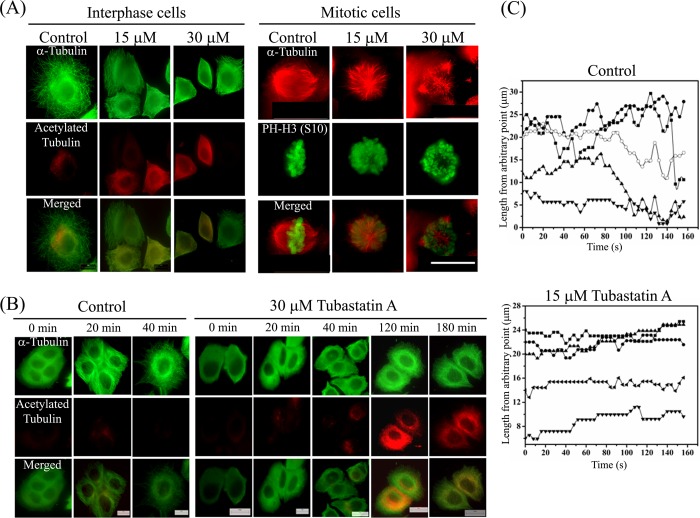
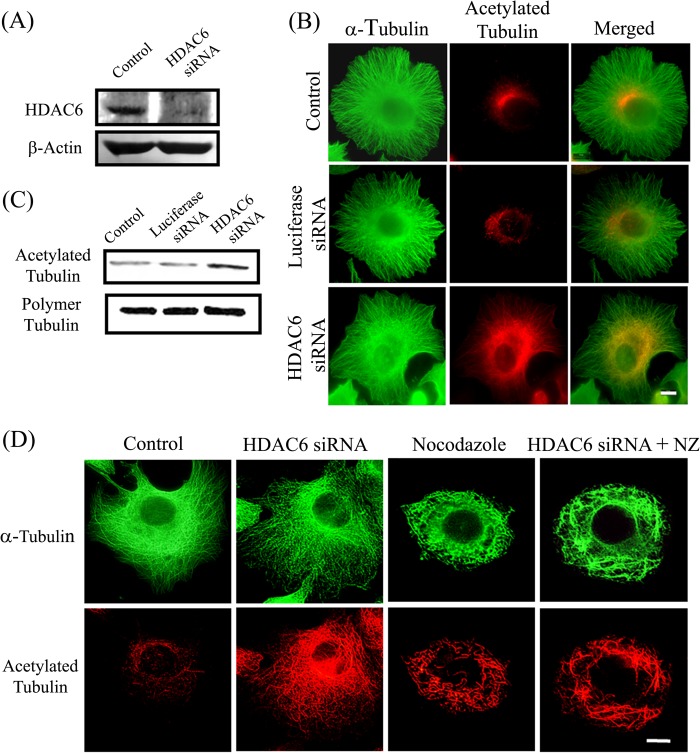
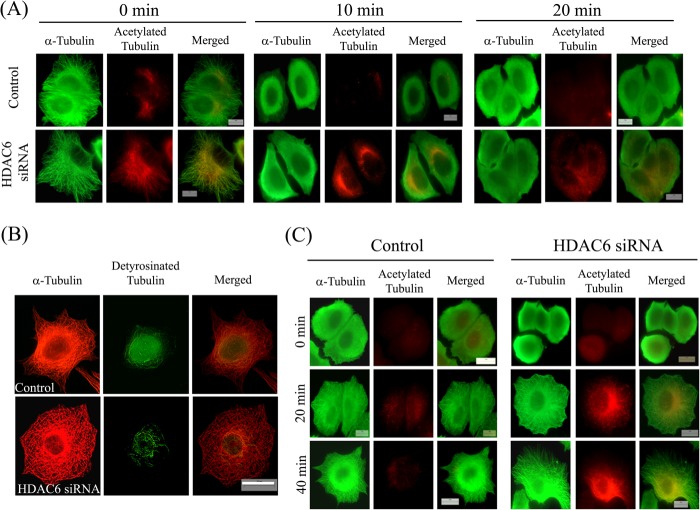
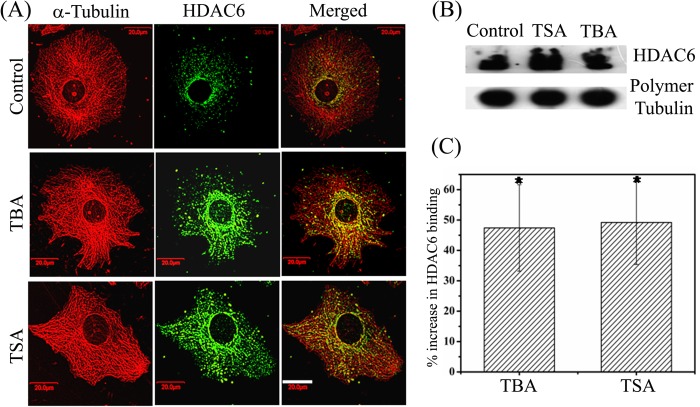
The post-translational modification of tubulin appears to be a highly controlled mechanism that regulates microtubule functioning. Acetylation of the ε-amino group of Lys-40 of α-tubulin marks stable microtubules, although the causal relationship between tubulin acetylation and microtubule stability has remained poorly understood. HDAC6, the tubulin deacetylase, plays a key role in maintaining typical distribution of acetylated microtubules in cells. Here, by using tubastatin A, an HDAC6-specific inhibitor, and siRNA-mediated depletion of HDAC6, we have explored whether tubulin acetylation has a role in regulating microtubule stability. We found that whereas both pharmacological inhibition of HDAC6 as well as its depletion enhance microtubule acetylation, only pharmacological inhibition of HDAC6 activity leads to an increase in microtubule stability against cold and nocodazole-induced depolymerizing conditions. Tubastatin A treatment suppressed the dynamics of individual microtubules in MCF-7 cells and delayed the reassembly of depolymerized microtubules. Interestingly, both the localization of HDAC6 on microtubules and the amount of HDAC6 associated with polymeric fraction of tubulin were found to increase in the tubastatin A-treated cells compared with the control cells, suggesting that the pharmacological inhibition of HDAC6 enhances the binding of HDAC6 to microtubules. The evidence presented in this study indicated that the increased binding of HDAC6, rather than the acetylation per se, causes microtubule stability. The results are in support of a hypothesis that in addition to its deacetylase function, HDAC6 might function as a MAP that regulates microtubule dynamics under certain conditions.
Keywords: Histone Deacetylase; Histone Deacetylase 6; Histone Deacetylase Inhibitors; MAPs; Microtubule Dynamics; Microtubules; Post-translational Modification; Tubastatin A; Tubulin Acetylation.
Figures
References
-
- Janke C., Bulinski J. C. (2011) Post-translational regulation of the microtubule cytoskeleton: mechanisms and functions. Nat. Rev. Mol. Cell Biol. 12, 773–786 - PubMed
-
- Hubbert C., Guardiola A., Shao R., Kawaguchi Y., Ito A., Nixon A., Yoshida M., Wang X. F., Yao T. P. (2002) HDAC6 is a microtubule-associated deacetylase. Nature 417, 455–458 - PubMed
-
- North B. J., Marshall B. L., Borra M. T., Denu J. M., Verdin E. (2003) The human Sir2 ortholog, SIRT2, is an NAD+-dependent tubulin deacetylase. Mol. Cell 11, 437–444 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
