

Distinctive profile of IsomiR expression and novel microRNAs in rat heart left ventricle
- PMID: 23799049
- PMCID: PMC3683050
- DOI: 10.1371/journal.pone.0065809
Distinctive profile of IsomiR expression and novel microRNAs in rat heart left ventricle
Abstract
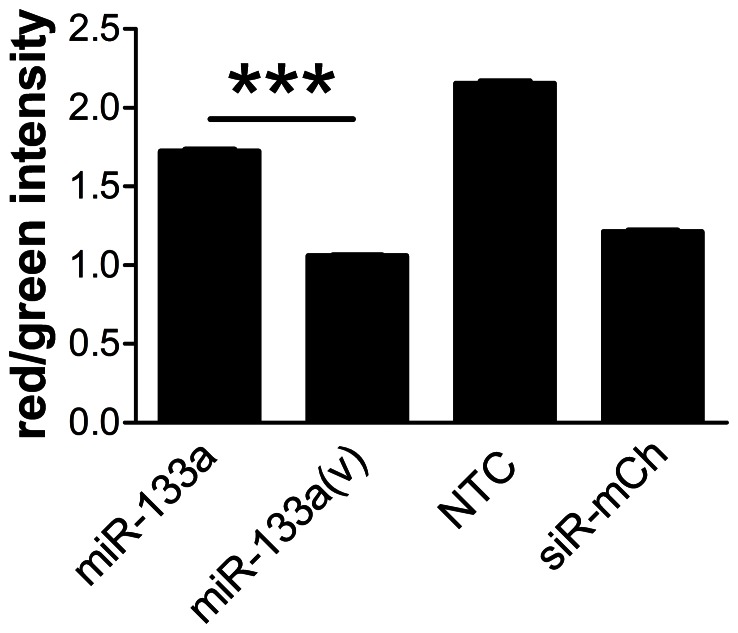
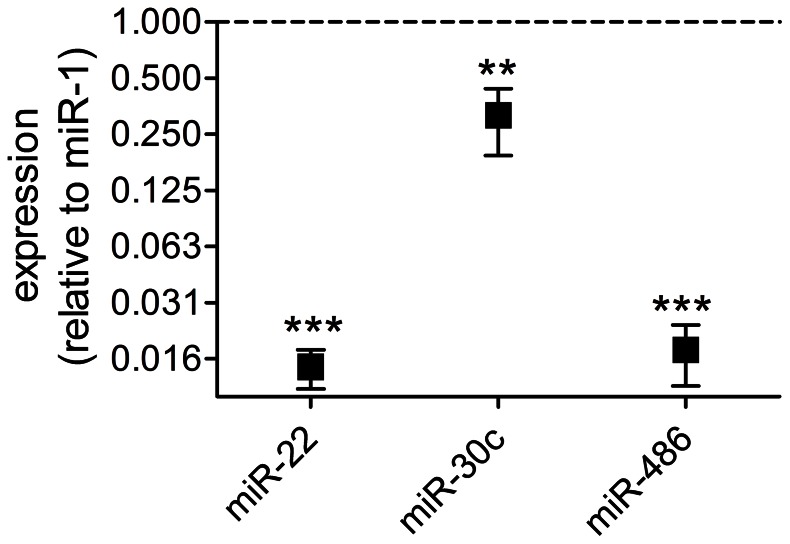
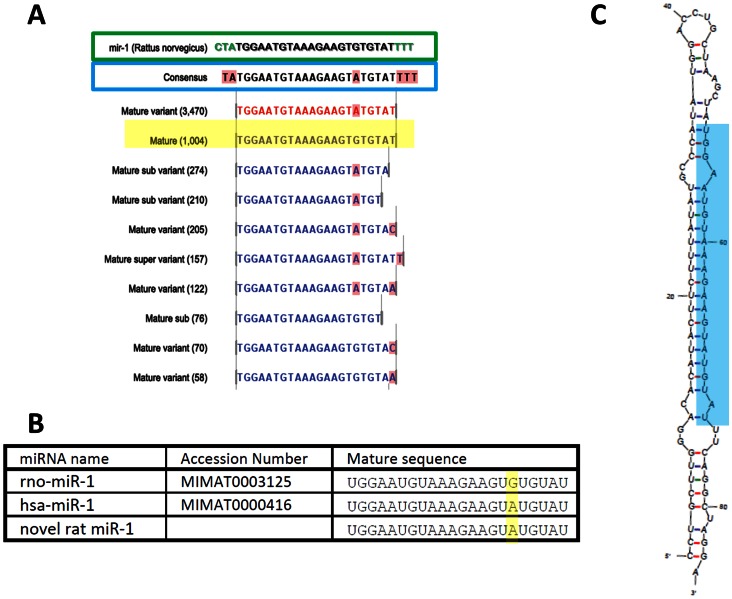
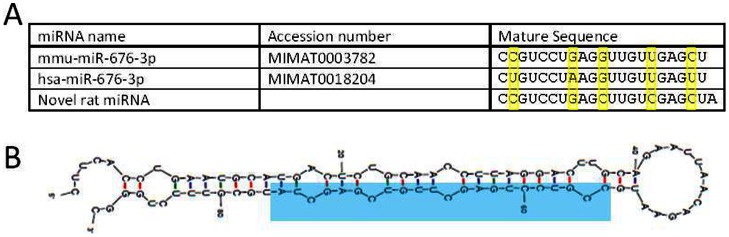
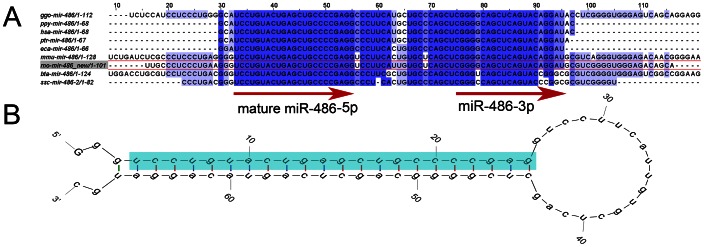
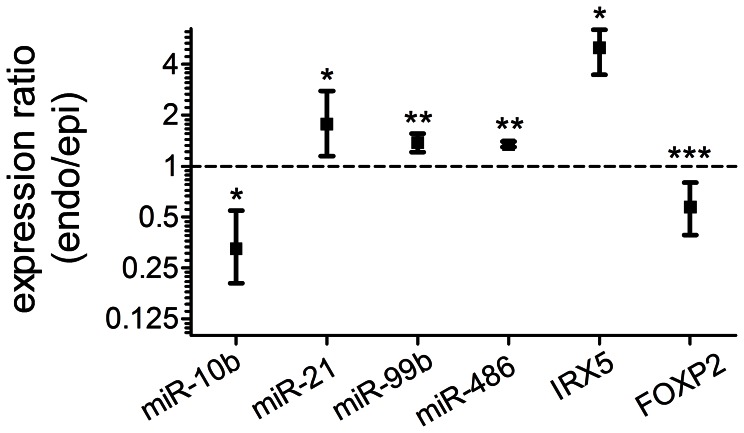
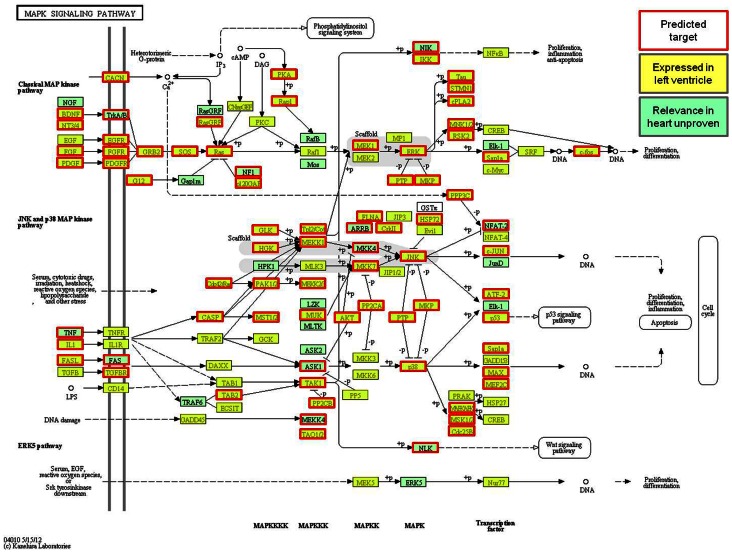
MicroRNAs (miRNAs) are single-stranded non-coding RNAs that negatively regulate target gene expression through mRNA cleavage or translational repression. There is mounting evidence that they play critical roles in heart disease. The expression of known miRNAs in the heart has been studied at length by microarray and quantitative PCR but it is becoming evident that microRNA isoforms (isomiRs) are potentially physiologically important. It is well known that left ventricular (patho)physiology is influenced by transmural heterogeneity of cardiomyocyte phenotype, and this likely reflects underlying heterogeneity of gene expression. Given the significant role of miRNAs in regulating gene expression, knowledge of how the miRNA profile varies across the ventricular wall will be crucial to better understand the mechanisms governing transmural physiological heterogeneity. To determinine miRNA/isomiR expression profiles in the rat heart we investigated tissue from different locations across the left ventricular wall using deep sequencing. We detected significant quantities of 145 known rat miRNAs and 68 potential novel orthologs of known miRNAs, in mature, mature* and isomiR formation. Many isomiRs were detected at a higher frequency than their canonical sequence in miRBase and have different predicted targets. The most common miR-133a isomiR was more effective at targeting a construct containing a sequence from the gelsolin gene than was canonical miR-133a, as determined by dual-fluorescence assay. We identified a novel rat miR-1 homolog from a second miR-1 gene; and a novel rat miRNA similar to miR-676. We also cloned and sequenced the rat miR-486 gene which is not in miRBase (v18). Signalling pathways predicted to be targeted by the most highly detected miRNAs include Ubiquitin-mediated Proteolysis, Mitogen-Activated Protein Kinase, Regulation of Actin Cytoskeleton, Wnt signalling, Calcium Signalling, Gap junctions and Arrhythmogenic Right Ventricular Cardiomyopathy. Most miRNAs are not expressed in a gradient across the ventricular wall, with exceptions including miR-10b, miR-21, miR-99b and miR-486.
Conflict of interest statement
Figures
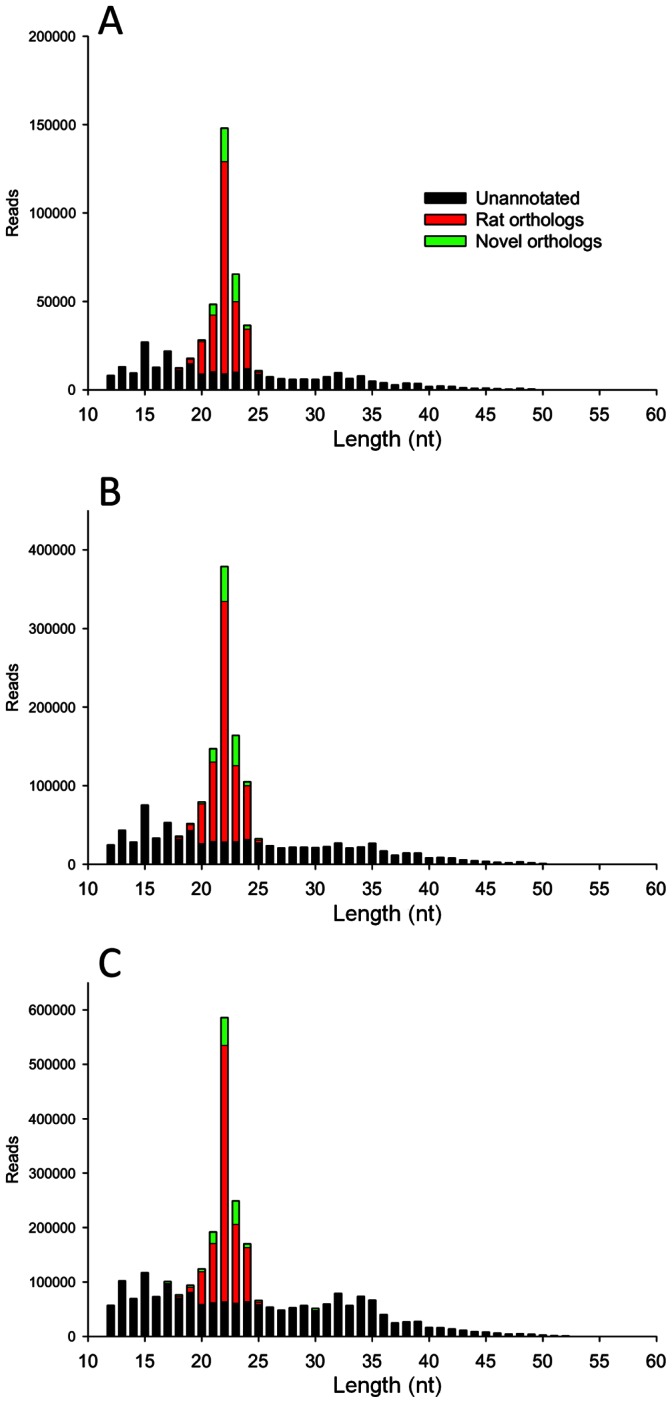
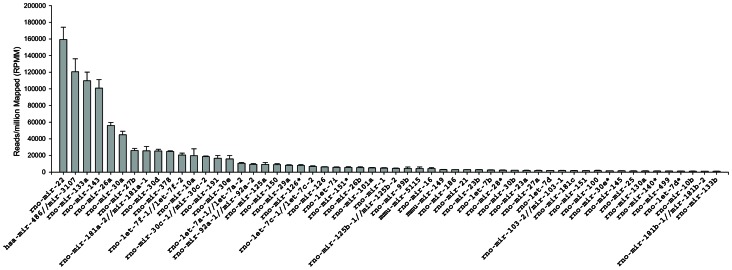
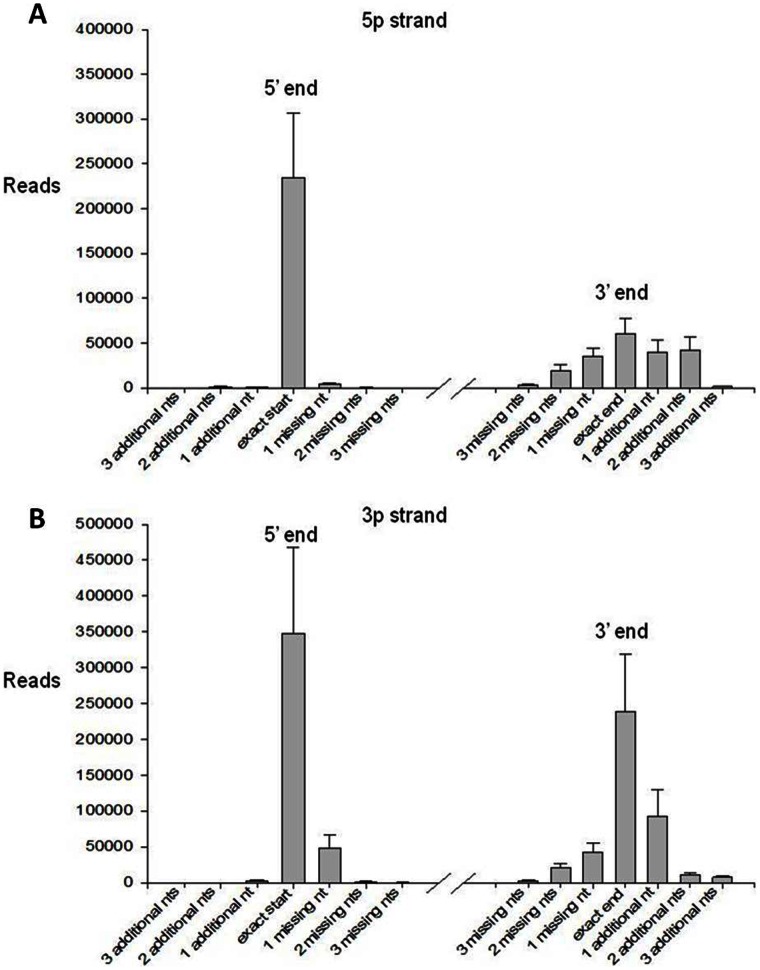
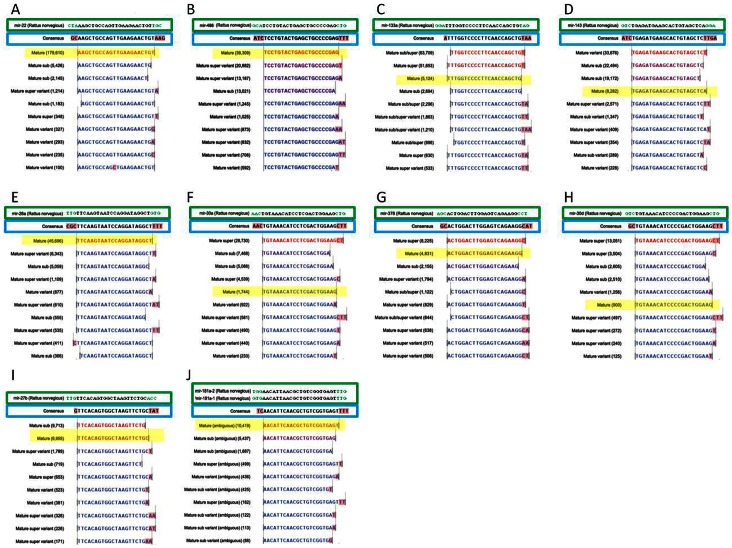
References
-
- Bartel DP (2004) MicroRNAs: genomics, biogenesis, mechanism, and function. Cell 116: 281–297. - PubMed
-
- D'Alessandra Y, Pompilio G, Capogrossi MC (2012) MicroRNAs and myocardial infarction. Curr Opin Cardiol 27: 228–235. - PubMed
-
- Bostjancic E, Zidar N, Stajer D, Glavac D (2010) MicroRNAs miR-1, miR-133a, miR-133b and miR-208 are dysregulated in human myocardial infarction. Cardiology 115: 163–169. - PubMed
-
- Gladka MM, da Costa Martins PA, De Windt LJ (2012) Small changes can make a big difference - microRNA regulation of cardiac hypertrophy. J Mol Cell Cardiol 52: 74–82. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
