

Different immunity elicited by recombinant H5N1 hemagglutinin proteins containing pauci-mannose, high-mannose, or complex type N-glycans
- PMID: 23799128
- PMCID: PMC3682957
- DOI: 10.1371/journal.pone.0066719
Different immunity elicited by recombinant H5N1 hemagglutinin proteins containing pauci-mannose, high-mannose, or complex type N-glycans
Abstract
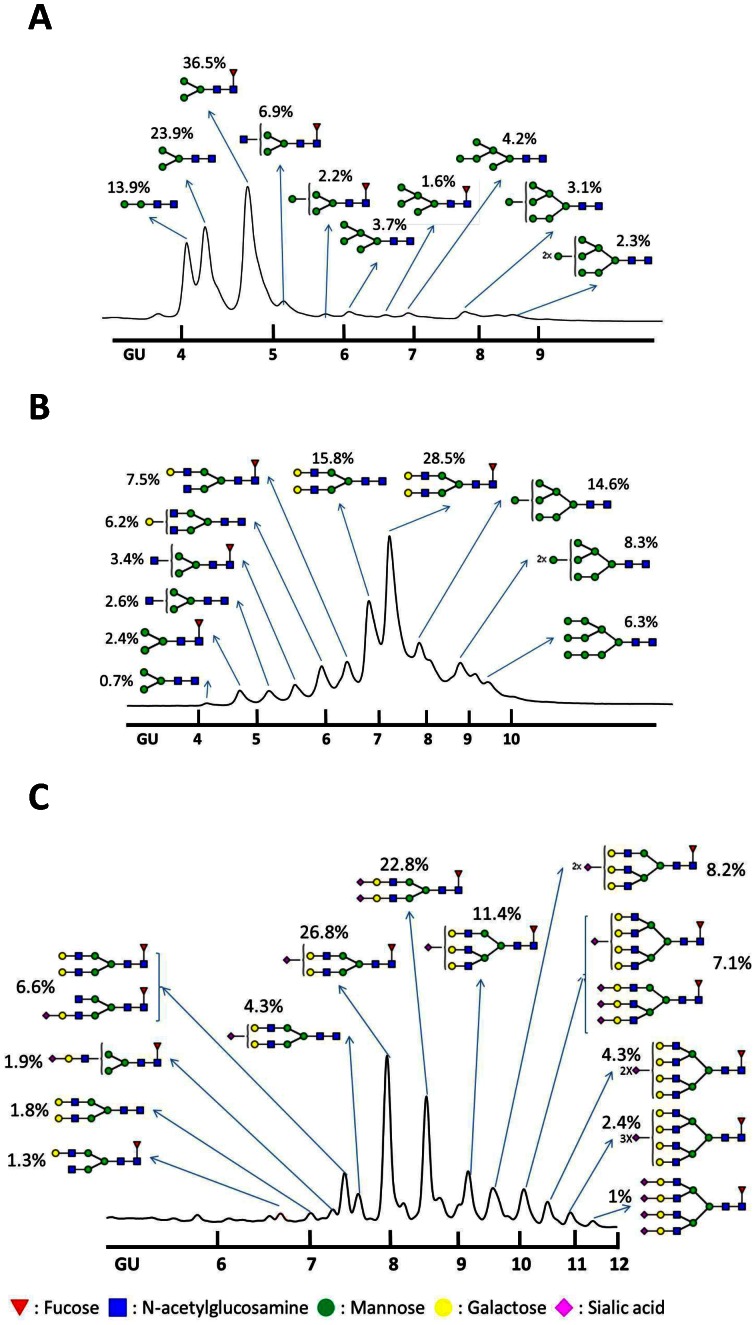
Highly pathogenic avian influenza H5N1 viruses can result in poultry and occasionally in human mortality. A safe and effective H5N1 vaccine is urgently needed to reduce the pandemic potential. Hemagglutinin (HA), a major envelope protein accounting for approximately 80% of spikes in influenza virus, is often used as a major antigen for subunit vaccine development. In this study, we conducted a systematic study of the immune response against influenza virus infection following immunization with recombinant HA proteins expressed in insect (Sf9) cells, insect cells that contain exogenous genes for elaborating N-linked glycans (Mimic) and mammalian cells (CHO). While the antibody titers are higher with the insect cell derived HA proteins, the neutralization and HA inhibition titers are much higher with the mammalian cell produced HA proteins. Recombinant HA proteins containing tri- or tetra-antennary complex, terminally sialylated and asialyated-galactose type N-glycans induced better protective immunity in mice to lethal challenge. The results are highly relevant to issues that should be considered in the production of fragment vaccines.
Conflict of interest statement
Figures
References
-
- World Health Organization. Cumulative number of confirmed human cases of avian influenza A(H5N1) reported to WHO. Available at: http://www.who.int/influenza/human_animal_interface/H5N1_cumulative_tabl.... Accessed 01 Jan 2013.
-
- Fouchier RA, Herfst S, Osterhaus AD (2012) Public health and biosecurity. Restricted data on influenza H5N1 virus transmission. Science 335: 662–663. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
