

Genetic variation in four maturity genes affects photoperiod insensitivity and PHYA-regulated post-flowering responses of soybean
- PMID: 23799885
- PMCID: PMC3698206
- DOI: 10.1186/1471-2229-13-91
Genetic variation in four maturity genes affects photoperiod insensitivity and PHYA-regulated post-flowering responses of soybean
Abstract
Background: Absence of or low sensitivity to photoperiod is necessary for short-day crops, such as rice and soybean, to adapt to high latitudes. Photoperiod insensitivity in soybeans is controlled by two genetic systems and involves three important maturity genes: E1, a repressor for two soybean orthologs of Arabidopsis FLOWERING LOCUS T (GmFT2a and GmFT5a), and E3 and E4, which are phytochrome A genes. To elucidate the diverse mechanisms underlying photoperiod insensitivity in soybean, we assessed the genotypes of four maturity genes (E1 through E4) in early-flowering photoperiod-insensitive cultivars and their association with post-flowering responses.
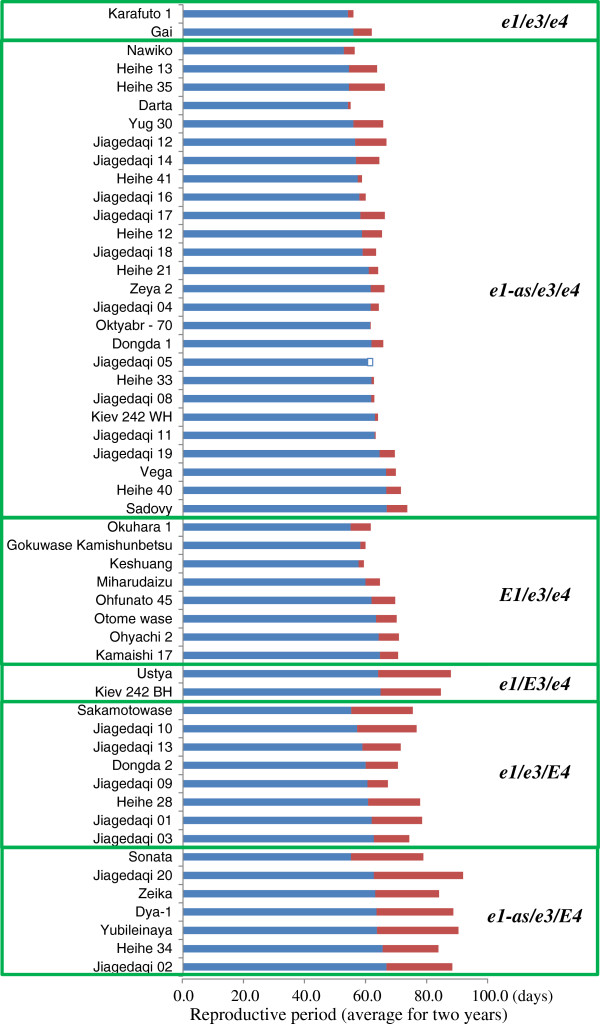
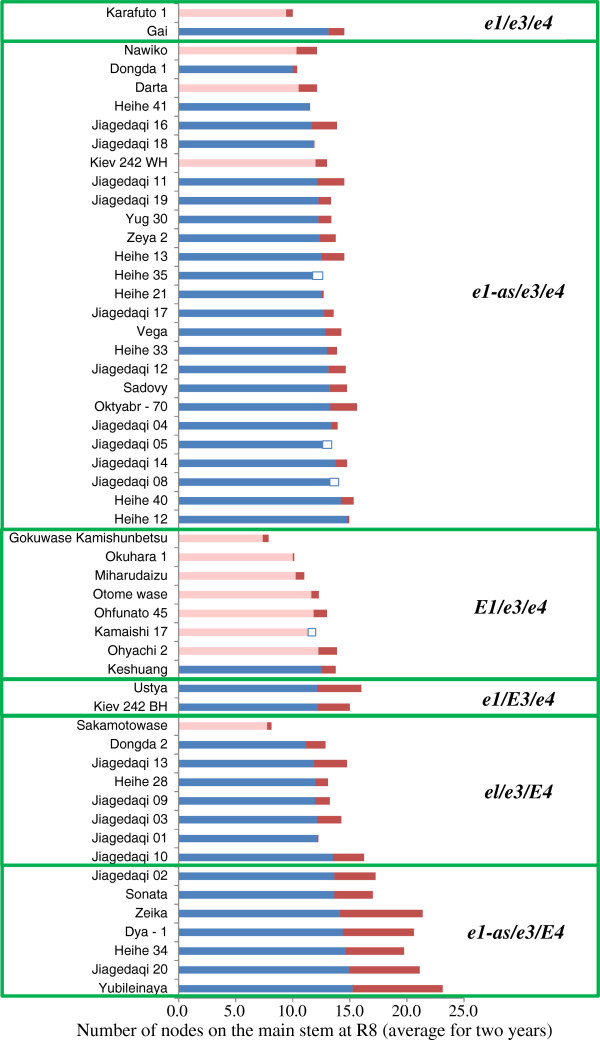
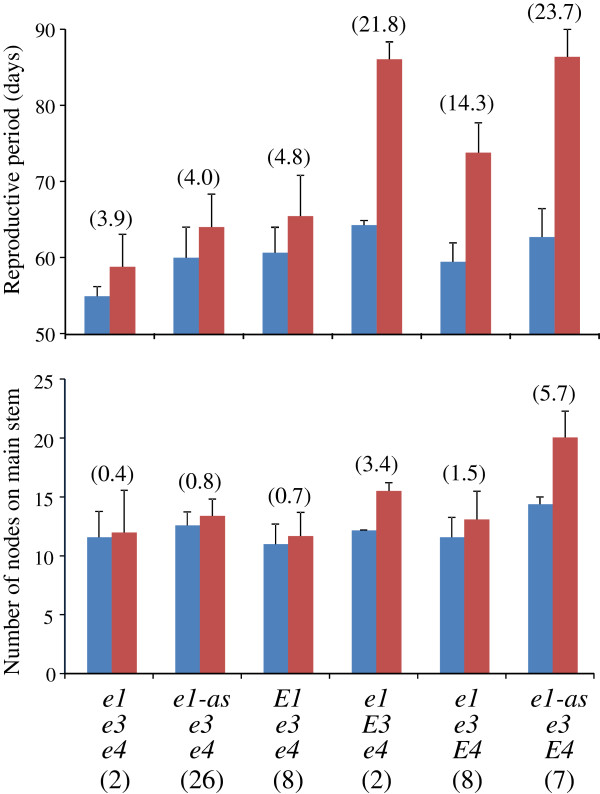
Results: We found two novel dysfunctional alleles in accessions originally considered to have a dominant E3 allele according to known DNA markers. The E3 locus, together with E1 and E4, contained multiple dysfunctional alleles. We identified 15 multi-locus genotypes, which we subdivided into 6 genotypic groups by classifying their alleles by function. Of these, the e1-as/e3/E4 genotypic group required an additional novel gene (different from E1, E3, and E4) to condition photoperiod insensitivity. Despite their common pre-flowering photoperiod insensitivity, accessions with different multi-locus genotypes responded differently to the post-flowering photoperiod. Cultivars carrying E3 or E4 were sensitive to photoperiod for post-flowering characteristics, such as reproductive period and stem growth after flowering. The phytochrome A-regulated expression of the determinate growth habit gene Dt1, an ortholog of Arabidopsis TERMINAL FLOWER1, was involved in the persistence of the vegetative activity at the stem apical meristem of flower-induced plants under long-day conditions.
Conclusions: Diverse genetic mechanisms underlie photoperiod insensitivity in soybean. At least three multi-locus genotypes consisting of various allelic combinations at E1, E3, and E4 conferred pre-flowering photoperiod insensitivity to soybean cultivars but led to different responses to photoperiod during post-flowering vegetative and reproductive development. The phyA genes E3 and E4 are major controllers underlying not only pre-flowering but also post-flowering photoperiod responses. The current findings improve our understanding of genetic diversity in pre-flowering photoperiod insensitivity and mechanisms of post-flowering photoperiod responses in soybean.
Figures



References
-
- Buzzell RI. Inheritance of a soybean flowering response to fluorescent-daylength conditions. Can J Genet Cytol. 1971;13:703–707.
-
- Buzzell RI, Voldeng HD. Inheritance of insensitivity to long day length. Soybean Genet Newsl. 1980;7:26–29.
-
- Saindon G, Voldeng HD, Beversdorf WD, Buzzell RI. Genetic control of long daylength response in soybean. Crop Sci. 1989;29:1436–1439. doi: 10.2135/cropsci1989.0011183X002900060021x. - DOI
-
- Cober ER, Tanner JW, Voldeng HD. Genetic control of photoperiod response in early-maturing near-isogenic soybean lines. Crop Sci. 1996;36:601–605. doi: 10.2135/cropsci1996.0011183X003600030013x. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
