

Review
doi: 10.1186/1741-7007-11-65.
Macromolecular juggling by ubiquitylation enzymes
Affiliations
- PMID: 23800009
- PMCID: PMC3748819
- DOI: 10.1186/1741-7007-11-65
Item in Clipboard
Review
Macromolecular juggling by ubiquitylation enzymes
BMC Biol.
.
Abstract
The posttranslational modification of target proteins with ubiquitin and ubiquitin-like proteins is accomplished by the sequential action of E1, E2, and E3 enzymes. Members of the E1 and E3 enzyme families can undergo particularly large conformational changes during their catalytic cycles, involving the remodeling of domain interfaces. This enables the efficient, directed and regulated handover of ubiquitin from one carrier to the next one. We review some of these conformational transformations, as revealed by crystallographic studies.
Figures
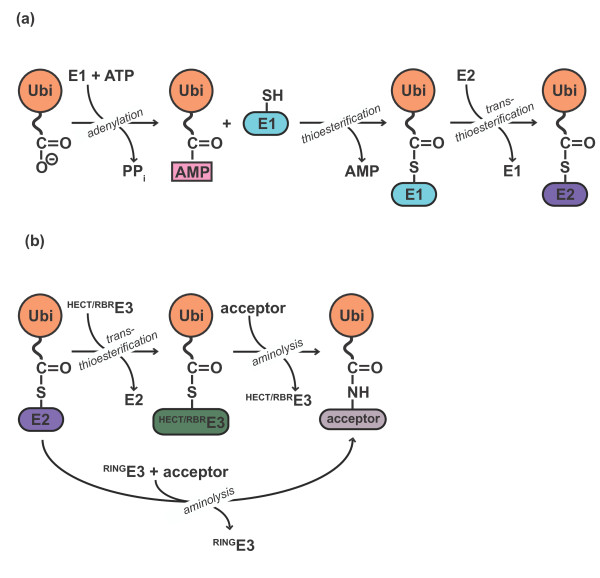
Ubiquitylation is a multistep reaction. (a) E1 enzymes use ATP to
activate the carboxyl terminus of ubiquitin (Ubi) as a high-energy anhydride
(Ubi-AMP). The E1 active site cysteine then attacks the adenylated ubiquitin
to form a thioester intermediate. Subsequently, the active site cysteine of
the E2 receives ubiquitin via trans-thioesterification. (b) E3
enzymes catalyze the formation of an isopeptide bond between the ubiquitin
carboxyl terminus and a primary amino group of an acceptor. The acceptor can
be a target protein (mono-ubiquitylation/ubiquitin chain initiation) or
another ubiquitin molecule (ubiquitin chain elongation). Catalysis by HECT-
and RBR-type E3 enzymes proceeds through an intermediate, in which the
ubiquitin carboxyl terminus is thioester-linked to a cysteine residue at the
active site of the E3, followed by aminolysis of the thioester. In contrast,
RING-type E3s catalyze direct transfer of ubiquitin from the E2 active site
cysteine to amino groups on the acceptor.
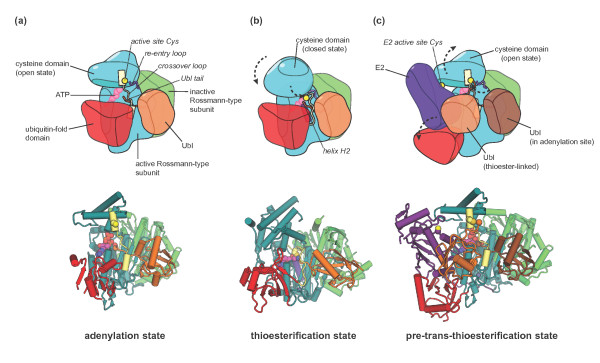
Conformational rearrangements in E1 enzymes. Cartoon representations
of distinct states in the catalytic cycle of canonical E1 enzymes.
(a) The adenylation state based on the crystal structure of
NAE1-UBA3 in complex with NEDD8 and ATP/Mg2+ [PDB: 1R4N] [32]. The carboxy-terminal tail of the Ubl is in the adenylation site
of the active Rossmann-type subunit of the E1, ready to nucleophilically
attack the α-phosphate of the ATP to form the Ubl-AMP intermediate. The
catalytic cysteine residue in the E1 cysteine domain is part of an
α-helix and is removed from the adenylation site, giving rise to an
open conformation of the cysteine domain. (b) The thioesterification
state as seen in a crystal structure of SAE1-UBA2 and SUMO covalently
coupled to an AMP analogue that mimics the tetrahedral intermediate
generated during thioesterification [PDB: 3KYD] [8]. Mediated by large conformational changes in the crossover and
re-entry loops, the cysteine domain is rotated with respect to the
Rossmann-type subunits. The helix containing the active site cysteine seen
in (a) has melted. In this closed conformation of the cysteine domain, the
catalytic cysteine nucleophile is in position to attack the adenylated
carboxyl terminus of the Ubl. The positive dipole of helix H2 in the active
Rossmann-type subunit (colored purple) is thought to favor this reaction [8]. (c) The trans-thioesterification state as represented by
a crystal structure of NAE1-UBA3 thioester-linked to NEDD8 and in complex
with an additional NEDD8 molecule, an E2 enzyme (Ubc12) and
ATP/Mg2+[35]. The cysteine domain of the E1 adopts an open orientation similar
to the adenylation state (a), but now holds the carboxyl terminus of the
thioester-linked Ubl close to the E2 active site (a Cys-to-Ala mutant of the
E2 was used in this study (see text)). The ubiquitin-fold domain has swung
away from its position in the previous states (a,b) to accommodate the E2
and contributes to E2 binding. In (a,c) domains found in NAE1-UBA3 but not
in SAE1-UBA2 were omitted for clarity. To see a rendition of a dynamic
transition between the structures shown in the lower panels of (a-c), see
Additional file 1. As noted in the movie legend,
the details of the trajectory linking individual structures is not realistic
and is simply meant to illustrate the nature of the conformational changes
rather than identify the nature of the transition pathway.
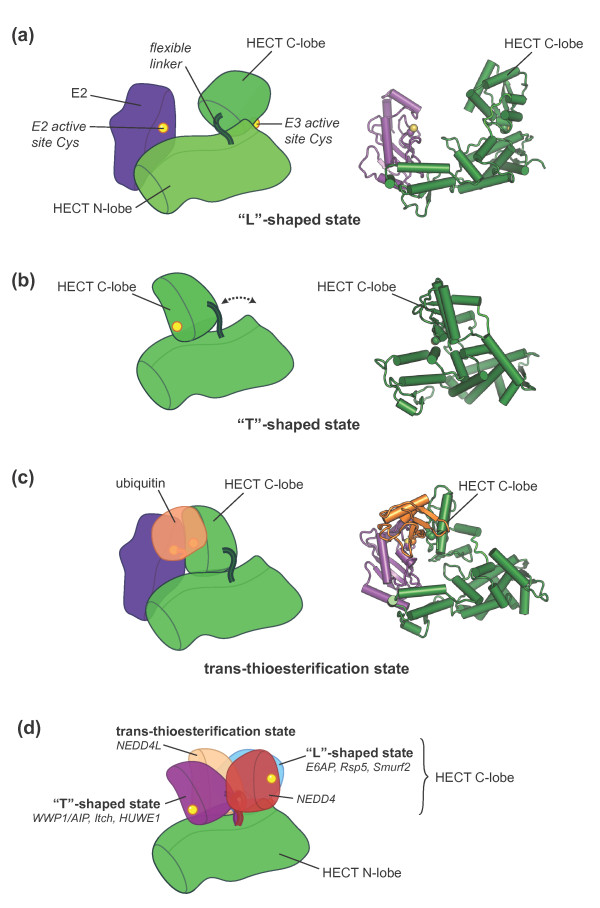
Swinging domains in HECT E3 enzymes. Cartoon representations of
crystal structures of various HECT domains. (a) Open,
‘L’-shaped conformation of E6AP (E3) in complex with UbcH7 (E2)
[PDB: 1C4Z] [62], (b) closed, ‘T’-shaped conformation of
WWP1/AIP [PDB: 1ND7] [63], and (c) trans-thioesterification complex of NEDD4L with a
ubiquitin-E2 (UbcH5B) conjugate [PDB:3JVZ] [64]. In (c) the E2 active site cysteine was mutated to serine
(colored yellow in our representation), resulting in an oxy-ester
linkage with ubiquitin in lieu of the native thioester. (d) Distinct
classes of C-lobe orientations based on the crystal structures of various
HECT domains (WWP1/AIP [PDB: 1ND7], Itch [PDB: 3TUG], HUWE1 [PDB: 3G1N,
3H1D], NEDD4L [PDB: 2ONI, 3JVZ], E6AP [PDB: 1C4Z], Rsp5 [PDB: 3OLM], Smurf2
[PDB: 1ZVD], NEDD [PDB: 2XBB]). A second unique C-lobe orientation observed
for NEDD [PDB: 2XBF] could not be displayed for clarity. In our
representation the HECT N-lobes are superimposed and only one of them is
displayed. Binding partners, such as E2 enzymes or ubiquitin, found in some
of the structures are not displayed.
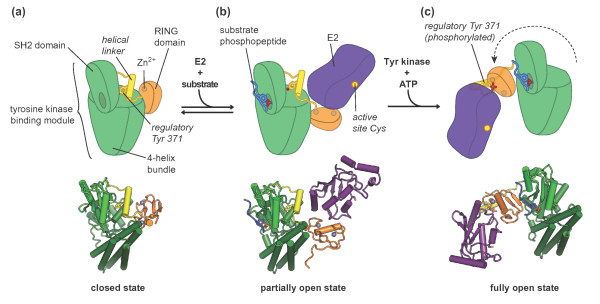
Regulatory rearrangements in Cbl proteins. (a) ‘Closed’
conformation of Cbl based on the crystal structure of the apo c-Cbl
amino-terminal region, comprising the tyrosine kinase binding module, the
helical linker region, and the RING domain [PDB: 2Y1M] [29]. The regulatory tyrosine, Y371, located in the helical linker
region, is buried in a hydrophobic core formed by the SH2 domain and the
four-helix bundle in the tyrosine kinase binding module. (b)
‘Partially open’ conformation of Cbl based on the co-crystal
structure of c-Cbl amino-terminal region with a ZAP70-derived phosphopeptide
and the E2 enzyme UbcH7 [PDB: 1FBV] [91]. Phosphopeptide binding induces a shift in the SH2 domain that
perturbs the interface between the helical linker and the tyrosine kinase
binding module, probably favoring dissociation of the RING domain from the
tyrosine kinase binding module and thus increasing the accessibility of the
E2 binding surface. (c) ‘Open’ conformation of Cbl based
on the co-crystal structure of phosphorylated c-Cbl bound to a ZAP7-derived
phosphopeptide and UbcH5B [PDB: 4A4C] [29]. The phosphorylated regulatory tyrosine, Tyr371, interacts with
residues in the E2 binding surface of the RING domain. The RING domain is
situated on the opposite side of the tyrosine kinase binding module compared
to (b).
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
