

Remodeling the integration of lipid metabolism between liver and adipose tissue by dietary methionine restriction in rats
- PMID: 23801581
- PMCID: PMC3781441
- DOI: 10.2337/db13-0501
Remodeling the integration of lipid metabolism between liver and adipose tissue by dietary methionine restriction in rats
Abstract
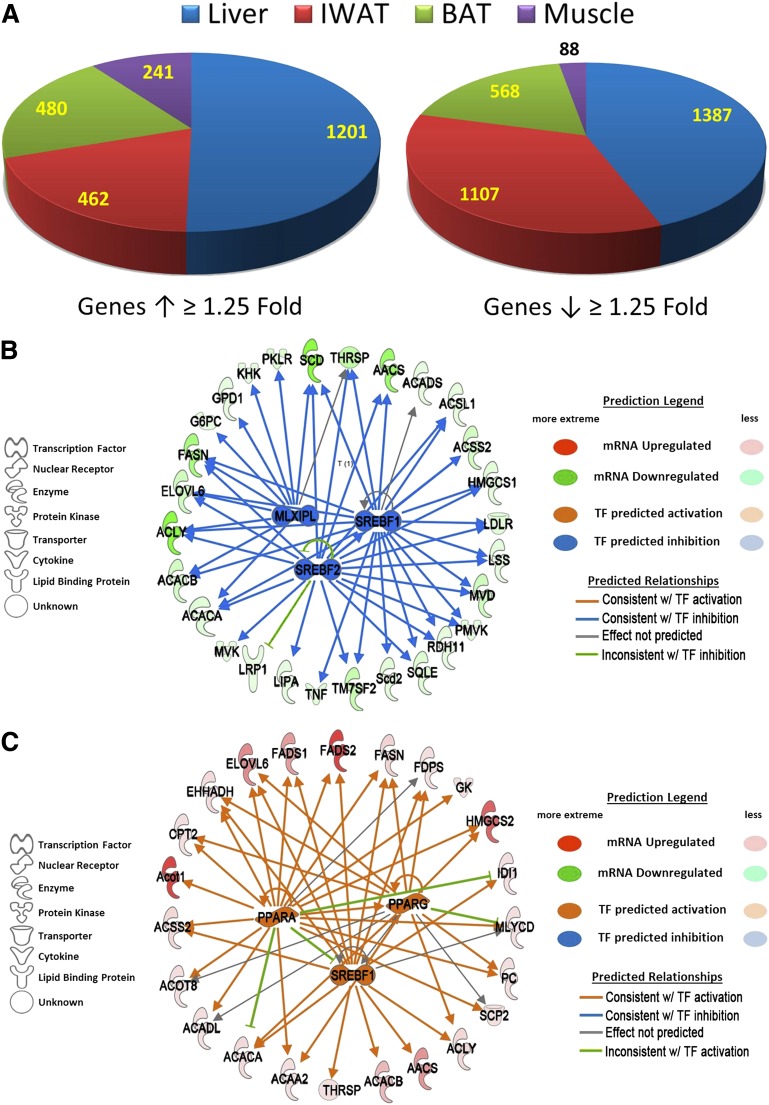
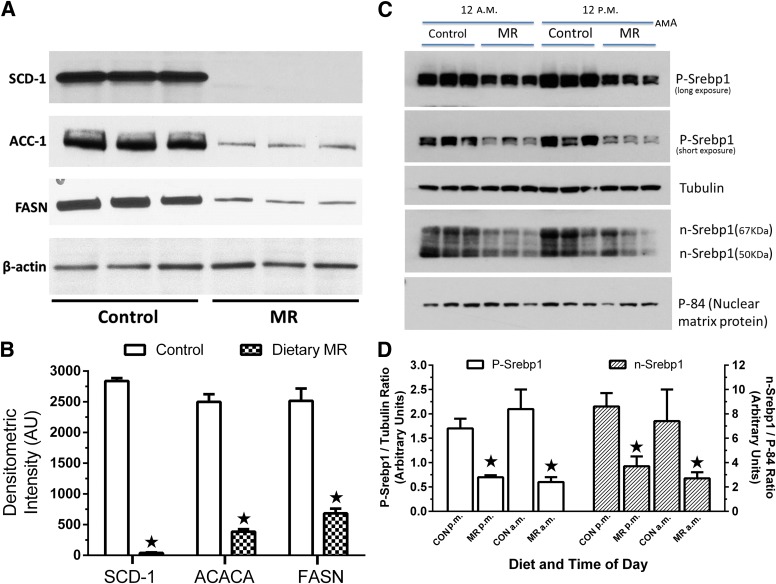
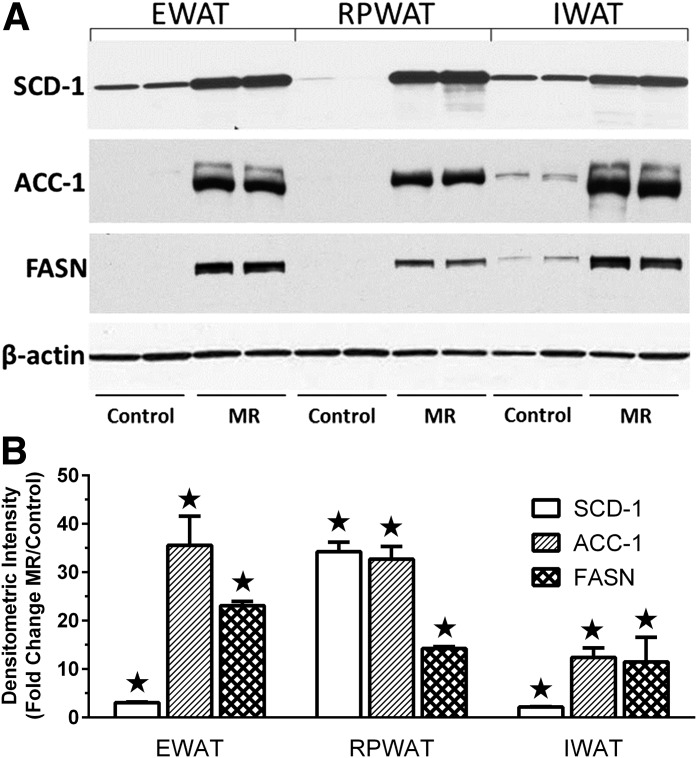
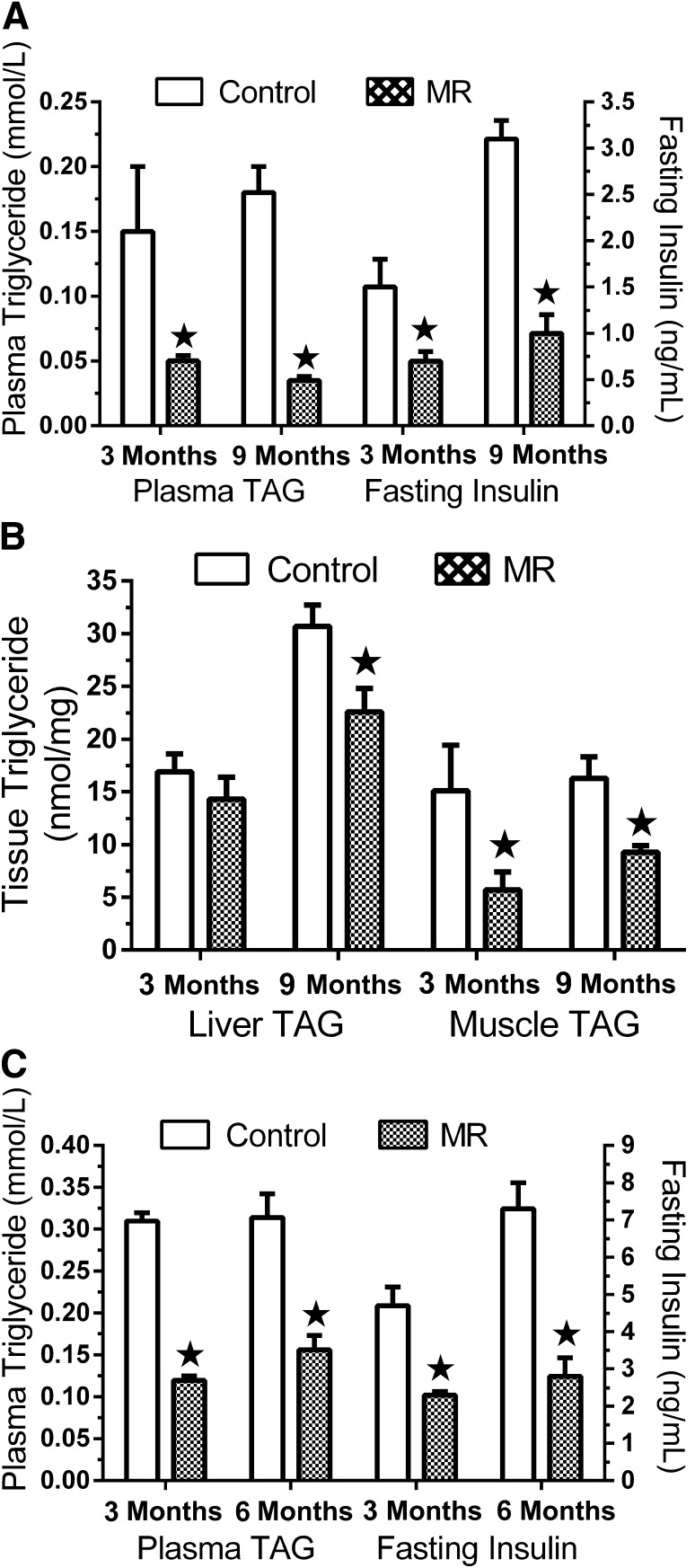
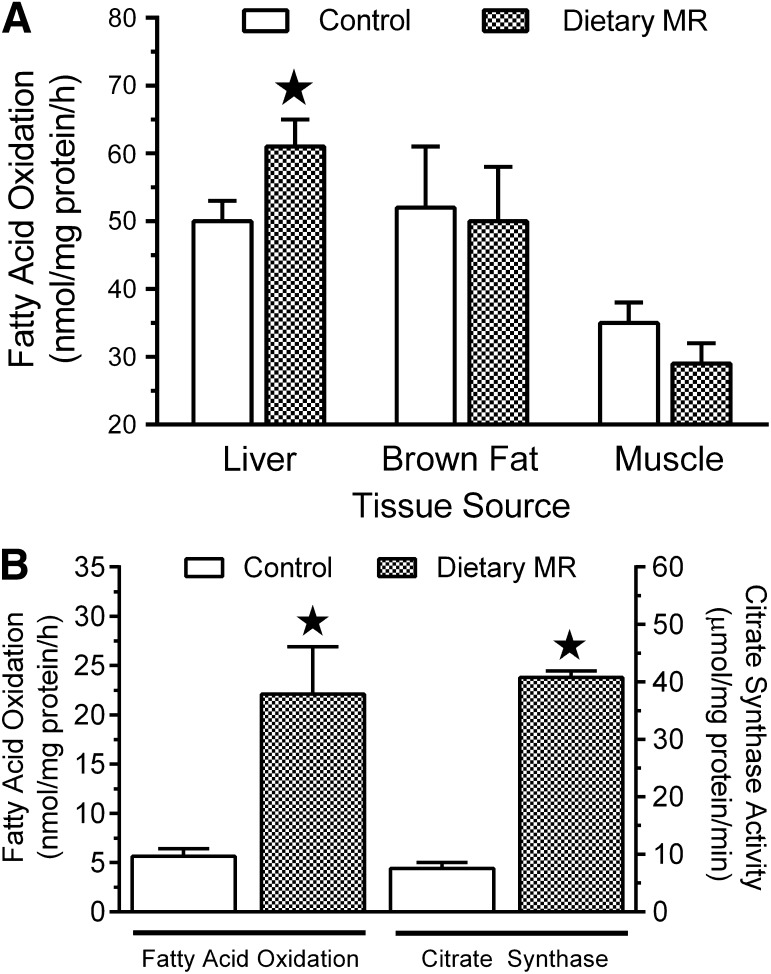
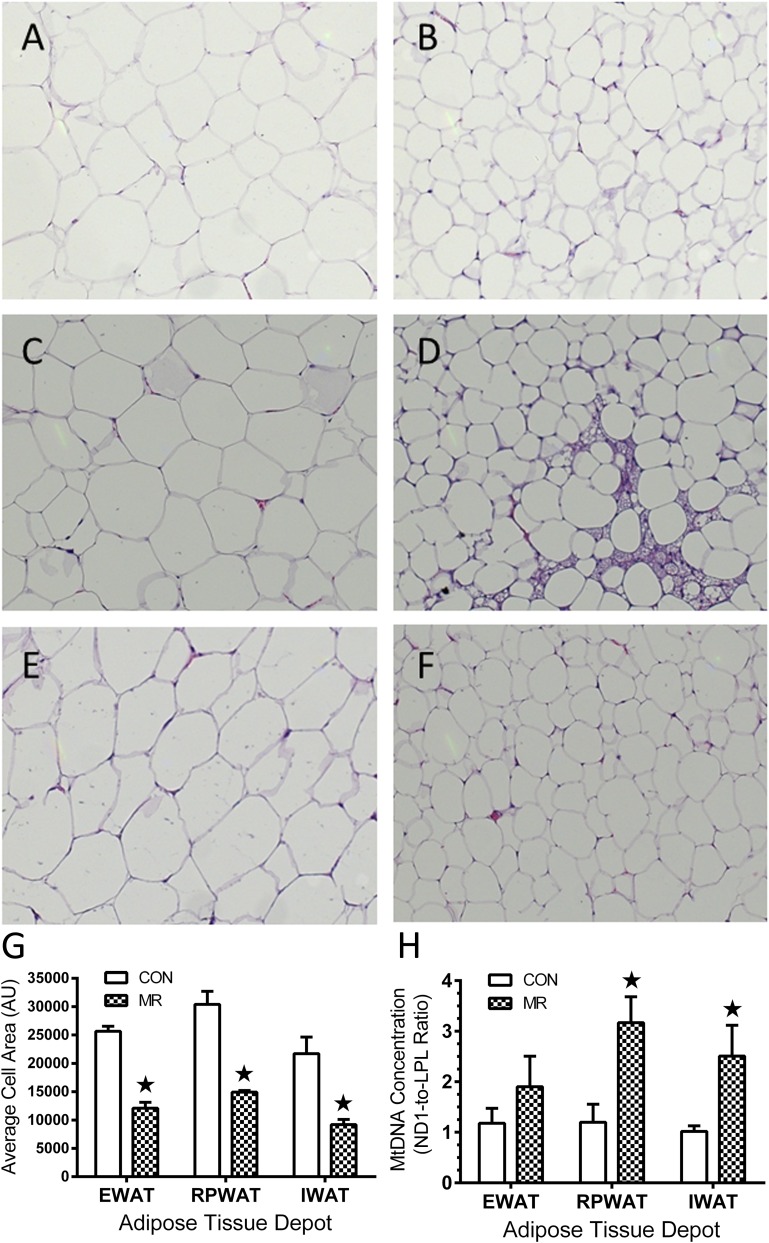
Dietary methionine restriction (MR) produces an integrated series of biochemical and physiological responses that improve biomarkers of metabolic health, limit fat accretion, and enhance insulin sensitivity. Using transcriptional profiling to guide tissue-specific evaluations of molecular responses to MR, we report that liver and adipose tissue are the primary targets of a transcriptional program that remodeled lipid metabolism in each tissue. The MR diet produced a coordinated downregulation of lipogenic genes in the liver, resulting in a corresponding reduction in the capacity of the liver to synthesize and export lipid. In contrast, the transcriptional response in white adipose tissue (WAT) involved a depot-specific induction of lipogenic and oxidative genes and a commensurate increase in capacity to synthesize and oxidize fatty acids. These responses were accompanied by a significant change in adipocyte morphology, with the MR diet reducing cell size and increasing mitochondrial density across all depots. The coordinated transcriptional remodeling of lipid metabolism between liver and WAT by dietary MR produced an overall reduction in circulating and tissue lipids and provides a potential mechanism for the increase in metabolic flexibility and enhanced insulin sensitivity produced by the diet.
Figures
References
-
- Gietzen DW. Neural mechanisms in the responses to amino acid deficiency. J Nutr 1993;123:610–625 - PubMed
-
- Gietzen DW, Erecius LF, Rogers QR. Neurochemical changes after imbalanced diets suggest a brain circuit mediating anorectic responses to amino acid deficiency in rats. J Nutr 1998;128:771–781 - PubMed
-
- Koehnle TJ, Russell MC, Gietzen DW. Rats rapidly reject diets deficient in essential amino acids. J Nutr 2003;133:2331–2335 - PubMed
-
- Guo F, Cavener DR. The GCN2 eIF2alpha kinase regulates fatty-acid homeostasis in the liver during deprivation of an essential amino acid. Cell Metab 2007;5:103–114 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U24 DK076169/DK/NIDDK NIH HHS/United States
- P20 RR021945/RR/NCRR NIH HHS/United States
- R01 DK096311/DK/NIDDK NIH HHS/United States
- P30 DK072476/DK/NIDDK NIH HHS/United States
- P20-GM-103528/GM/NIGMS NIH HHS/United States
- R01 DK-096311/DK/NIDDK NIH HHS/United States
- R21 DK-088319/DK/NIDDK NIH HHS/United States
- U24 DK-076169/DK/NIDDK NIH HHS/United States
- T32 DK-064584/DK/NIDDK NIH HHS/United States
- P60 DK020541/DK/NIDDK NIH HHS/United States
- P20 GM103528/GM/NIGMS NIH HHS/United States
- P30 GM118430/GM/NIGMS NIH HHS/United States
- R21 DK088319/DK/NIDDK NIH HHS/United States
- T32 DK064584/DK/NIDDK NIH HHS/United States
- P30 DK-072476/DK/NIDDK NIH HHS/United States
- R01 DK098439/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
