

Potent inhibition of DOT1L as treatment of MLL-fusion leukemia
- PMID: 23801631
- PMCID: PMC3739029
- DOI: 10.1182/blood-2013-04-497644
Potent inhibition of DOT1L as treatment of MLL-fusion leukemia
Abstract
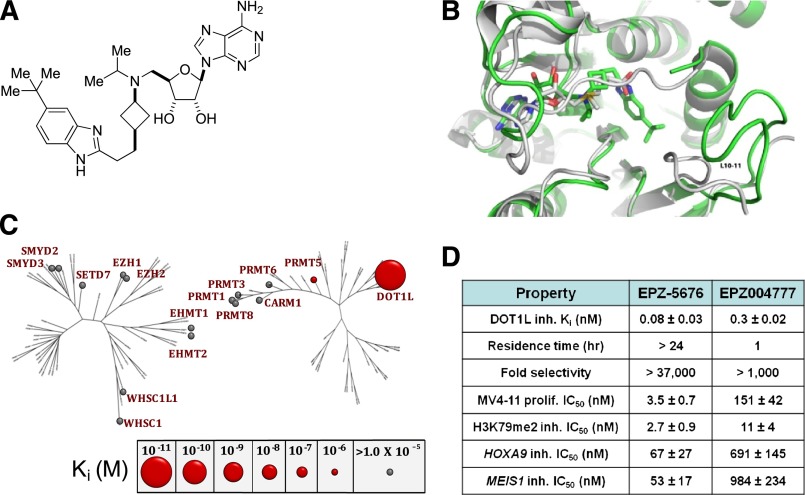
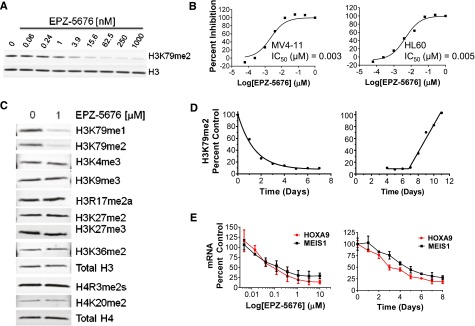
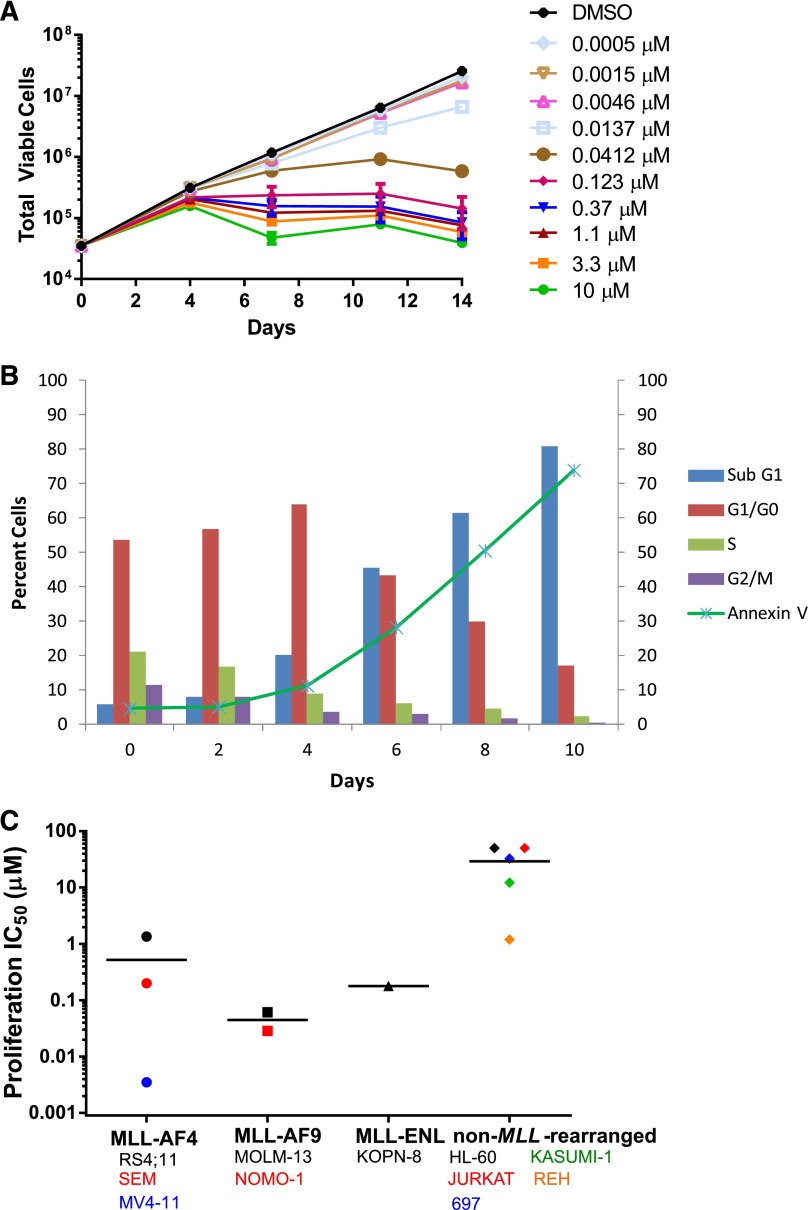
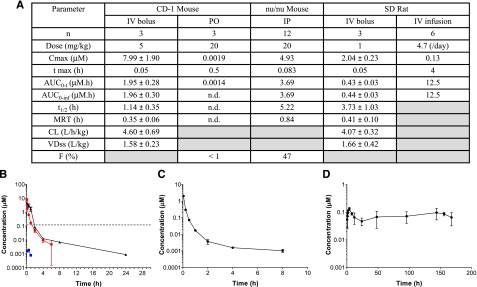
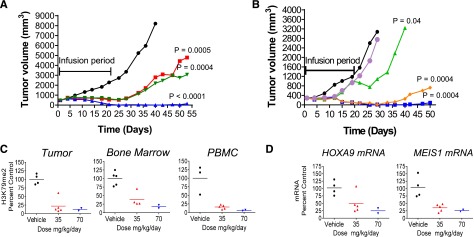
Rearrangements of the MLL gene define a genetically distinct subset of acute leukemias with poor prognosis. Current treatment options are of limited effectiveness; thus, there is a pressing need for new therapies for this disease. Genetic and small molecule inhibitor studies have demonstrated that the histone methyltransferase DOT1L is required for the development and maintenance of MLL-rearranged leukemia in model systems. Here we describe the characterization of EPZ-5676, a potent and selective aminonucleoside inhibitor of DOT1L histone methyltransferase activity. The compound has an inhibition constant value of 80 pM, and demonstrates 37 000-fold selectivity over all other methyltransferases tested. In cellular studies, EPZ-5676 inhibited H3K79 methylation and MLL-fusion target gene expression and demonstrated potent cell killing that was selective for acute leukemia lines bearing MLL translocations. Continuous IV infusion of EPZ-5676 in a rat xenograft model of MLL-rearranged leukemia caused complete tumor regressions that were sustained well beyond the compound infusion period with no significant weight loss or signs of toxicity. EPZ-5676 is therefore a potential treatment of MLL-rearranged leukemia and is under clinical investigation.
Figures
References
-
- Hess JL. MLL: a histone methyltransferase disrupted in leukemia. Trends Mol Med. 2004;10(10):500–507. - PubMed
-
- Krivtsov AV, Armstrong SA. MLL translocations, histone modifications and leukaemia stem-cell development. Nat Rev Cancer. 2007;7(11):823–833. - PubMed
-
- Tamai H, Inokuchi K. 11q23/MLL acute leukemia: update of clinical aspects. J Clin Exp Hematop. 2010;50(2):91–98. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
