

Morphogen gradients in development: from form to function
- PMID: 23801664
- PMCID: PMC3957335
- DOI: 10.1002/wdev.2
Morphogen gradients in development: from form to function
Abstract
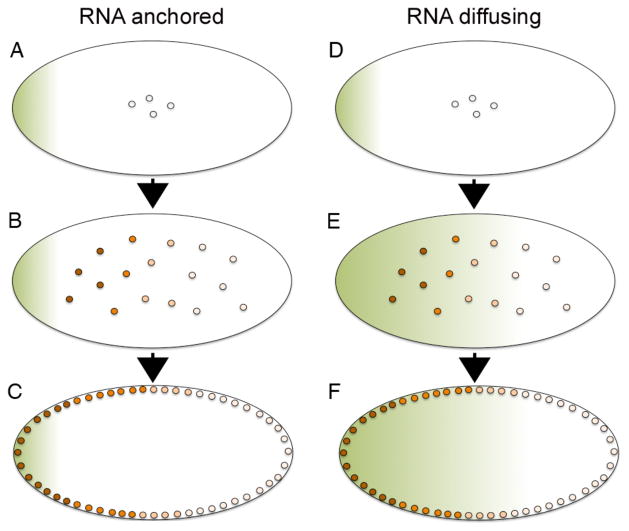
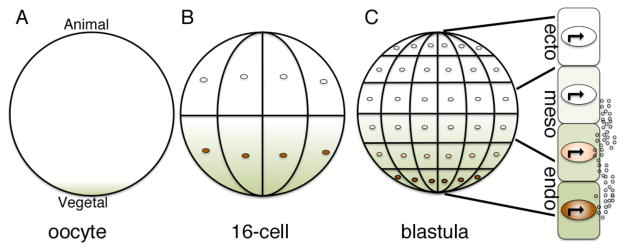
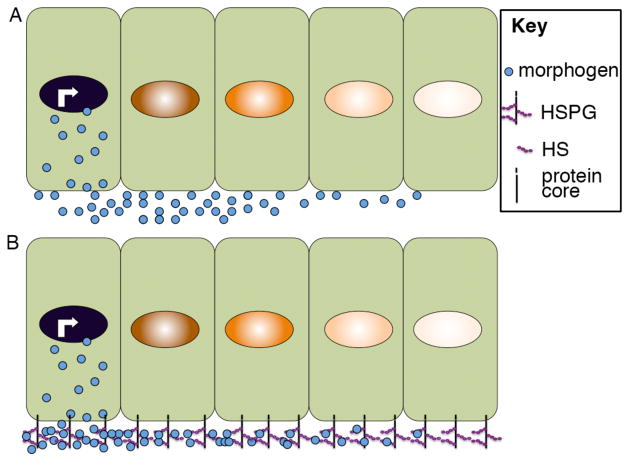
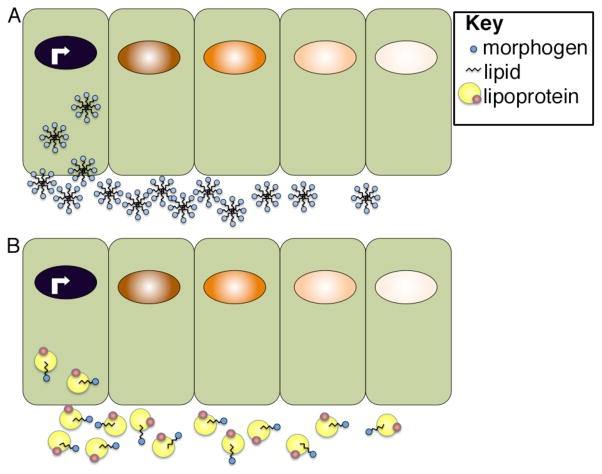
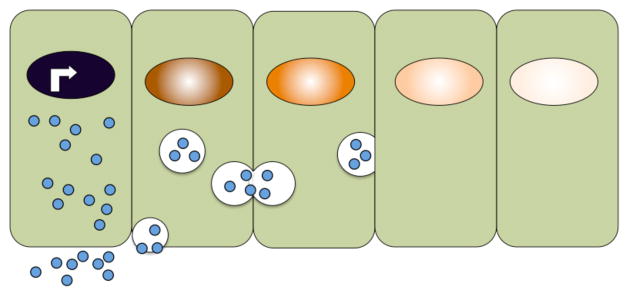
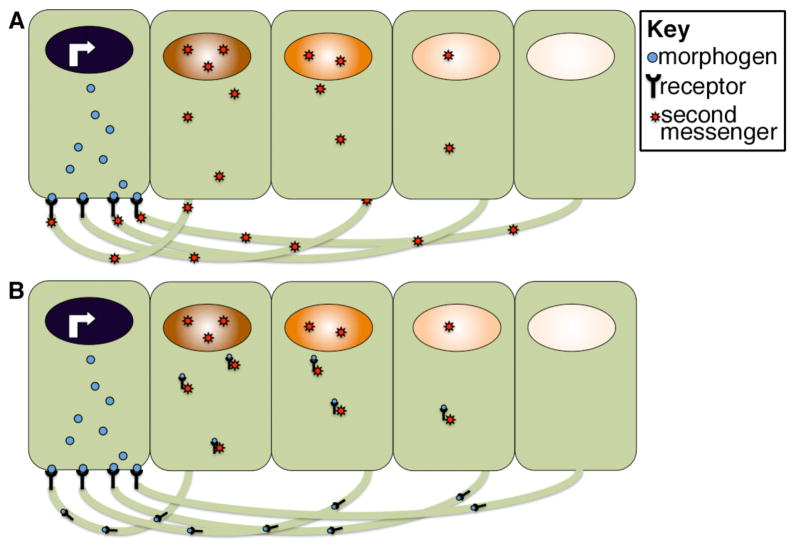
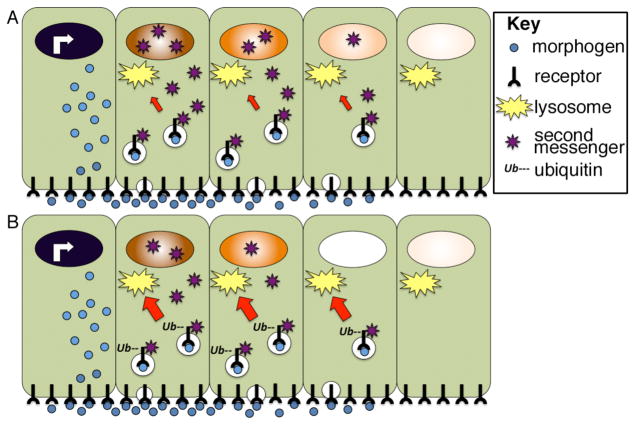
Morphogens are substances that establish a graded distribution and elicit distinct cellular responses in a dose-dependent manner. They function to provide individual cells within a field with positional information, which is interpreted to give rise to spatial patterns. Morphogens can consist of intracellular factors that set up a concentration gradient by diffusion in the cytoplasm. More commonly, morphogens comprise secreted proteins that form an extracellular gradient across a field of cells. Experimental studies and computational analyses have provided support for a number of diverse strategies by which extracellular morphogen gradients are formed. These include free diffusion in the extracellular space, restricted diffusion aided by interactions with heparan sulfate proteoglycans, transport on lipid-containing carriers or transport aided by soluble binding partners. More specialized modes of transport have also been postulated such as transcytosis, in which repeated rounds of secretion, endocytosis, and intracellular trafficking move morphogens through cells rather than around them, or cytonemes, which consist of filopodial extensions from signal-receiving cells that are hypothesized to reach out to morphogen-sending cells. Once the gradient has formed, cells must distinguish small differences in morphogen concentration and store this information even after the gradient has dissipated. This is often achieved by translating ligand concentration into a proportional increase in numbers of activated cell surface receptors that are internalized and continue to signal from endosomal compartments. Ultimately, this leads to activation of one or a few transcription factors that transduce this information into qualitatively distinct gene responses inside the nucleus.
Copyright © 2011 Wiley Periodicals, Inc.
Figures

References
Further Reading/Resources
-
- Gilbert . Developmental Biology. 9. 2010.
-
- The interactive fly. http://www.sdbonline.org/fly/aimain/1aahome.htm.
-
- Port F, Basler K. Wnt trafficking: new insights into Wnt maturation, secretion and spreading. Traffic. 2010;11:1265–1271. - PubMed
-
- Gallet A. Hedgehog morphogen: from secretion to reception. Trends Cell Biol. 2011;21:238–246. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
