

Comparisons of the embryonic development of Drosophila, Nasonia, and Tribolium
- PMID: 23801665
- PMCID: PMC5323069
- DOI: 10.1002/wdev.3
Comparisons of the embryonic development of Drosophila, Nasonia, and Tribolium
Abstract
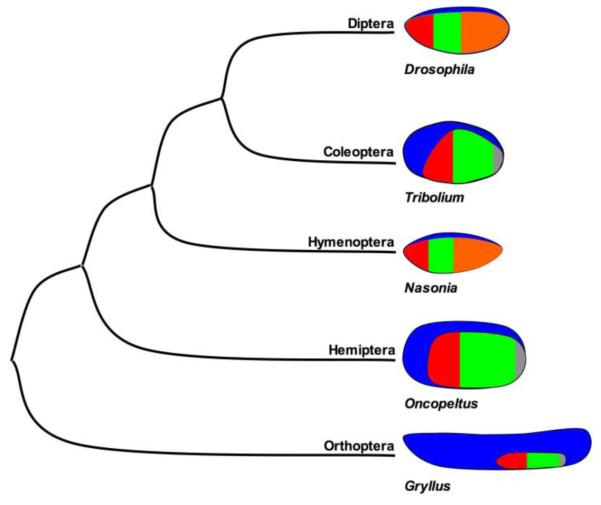
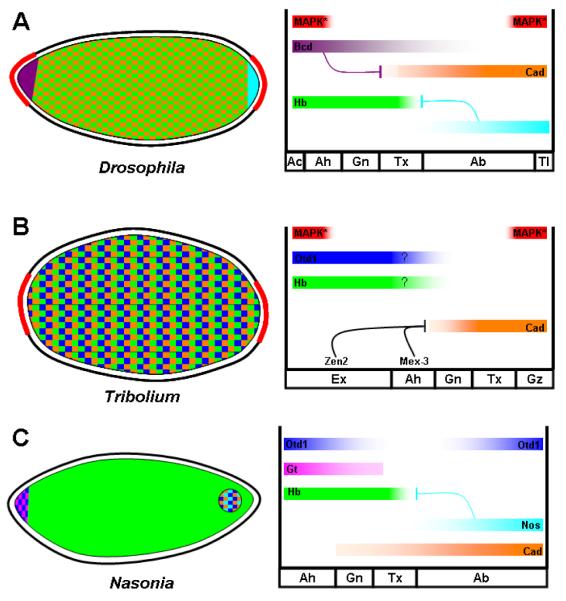
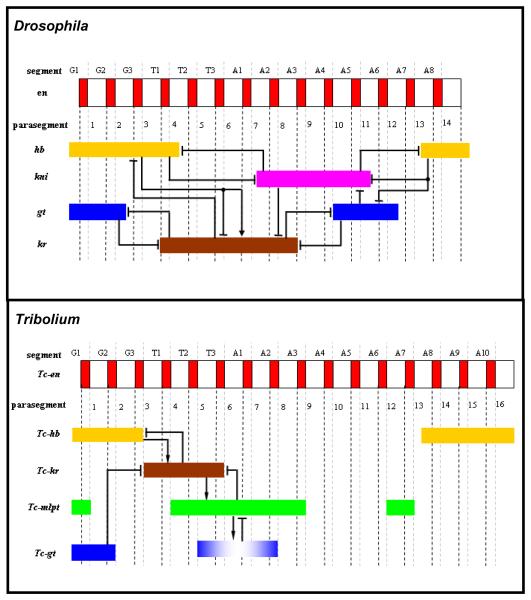
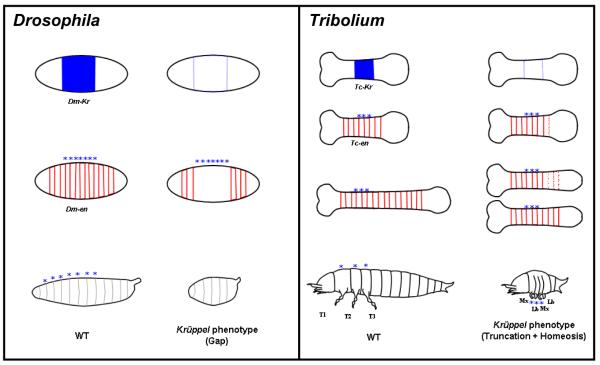
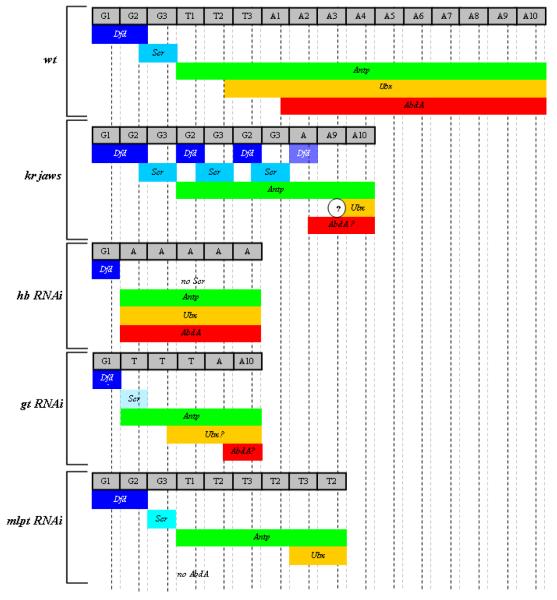
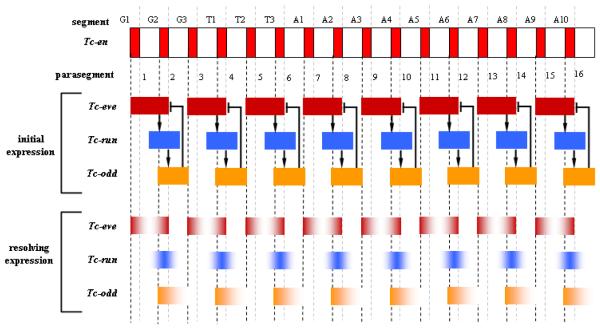
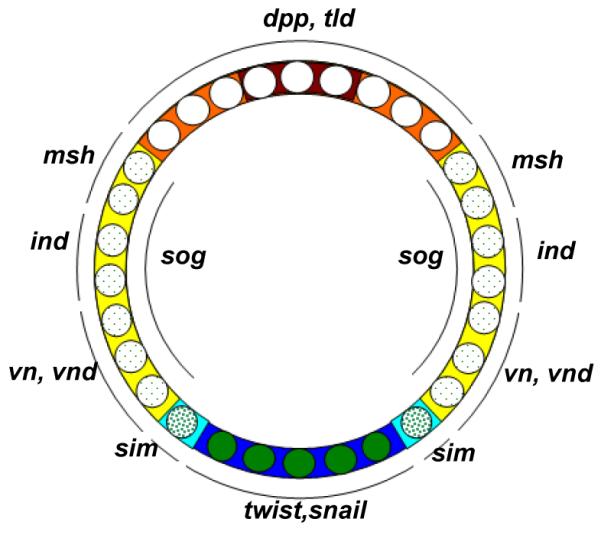
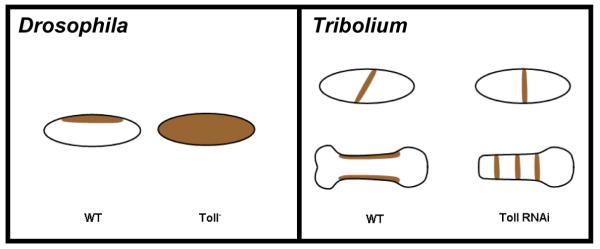
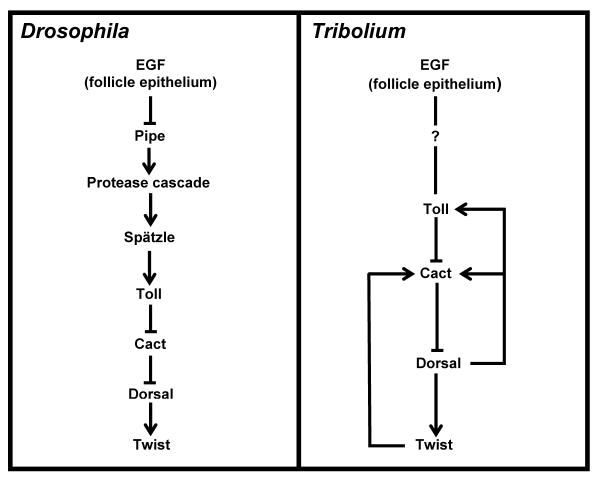
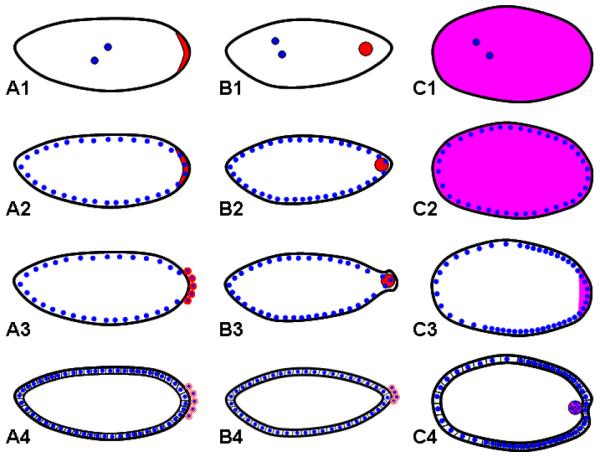
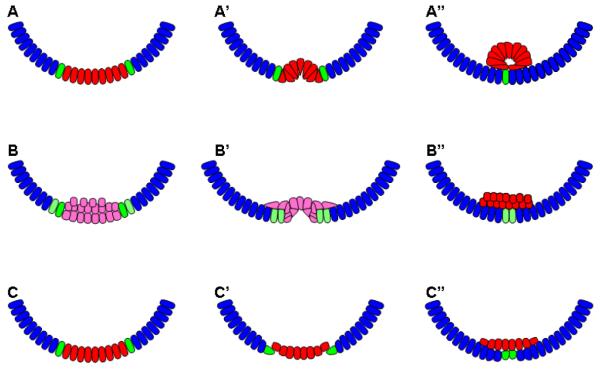
Studying the embryogenesis of diverse insect species is crucial to understanding insect evolution. Here, we review current advances in understanding the development of two emerging model organisms: the wasp Nasonia vitripennis and the beetle Tribolium castaneum in comparison with the well-studied fruit fly Drosophila melanogaster. Although Nasonia represents the most basally branching order of holometabolous insects, it employs a derived long germband mode of embryogenesis, more like that of Drosophila, whereas Tribolium undergoes an intermediate germband mode of embryogenesis, which is more similar to the ancestral mechanism. Comparing the embryonic development and genetic regulation of early patterning events in these three insects has given invaluable insights into insect evolution. The similar mode of embryogenesis of Drosophila and Nasonia is reflected in their reliance on maternal morphogenetic gradients. However, they employ different genes as maternal factors, reflecting the evolutionary distance separating them. Tribolium, on the other hand, relies heavily on self-regulatory mechanisms other than maternal cues, reflecting its sequential nature of segmentation and the need for reiterated patterning.
Copyright © 2011 Wiley Periodicals, Inc.
Figures
References
-
- Davis GK, Patel NH. Short, long, and beyond: Molecular and embryological approaches to insect segmentation. Annual Review of Entomology. 2002;47:669–699. - PubMed
-
- Tautz D, Friedrich M, Schröder R. Insect embryogenesis - what is ancestral and what is derived? Development. 1994;(Supplement):193–199.
-
- Richards S, Gibbs RA, Weinstock GM, Brown SJ, Denell R, Beeman RW, Gibbs R, Bucher G, Friedrich M, Grimmelikhuijzen CJP, et al. The genome of the model beetle and pest Tribolium castaneum. Nature. 2008;452:949–955. - PubMed
-
- Bucher G, Scholten J, Klingler M. Parental RNAi in Tribolium (Coleoptera) Curr Biol. 2002;12:R85–86. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
