

Diversification through multitrait evolution in a coevolving interaction
- PMID: 23801764
- PMCID: PMC3710874
- DOI: 10.1073/pnas.1307451110
Diversification through multitrait evolution in a coevolving interaction
Abstract
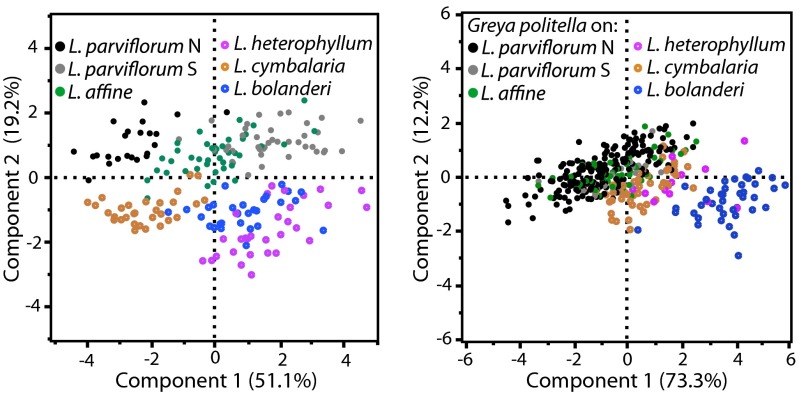
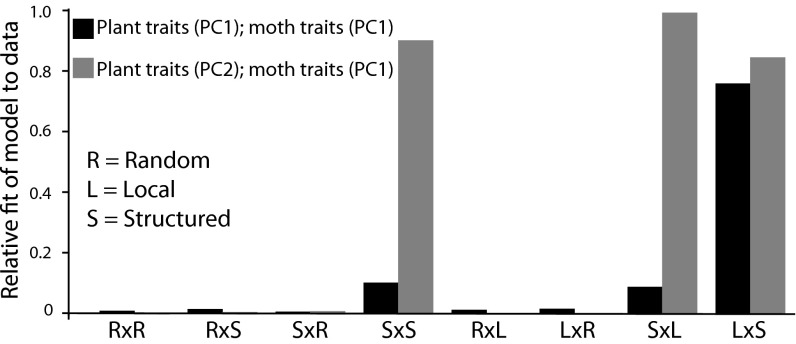
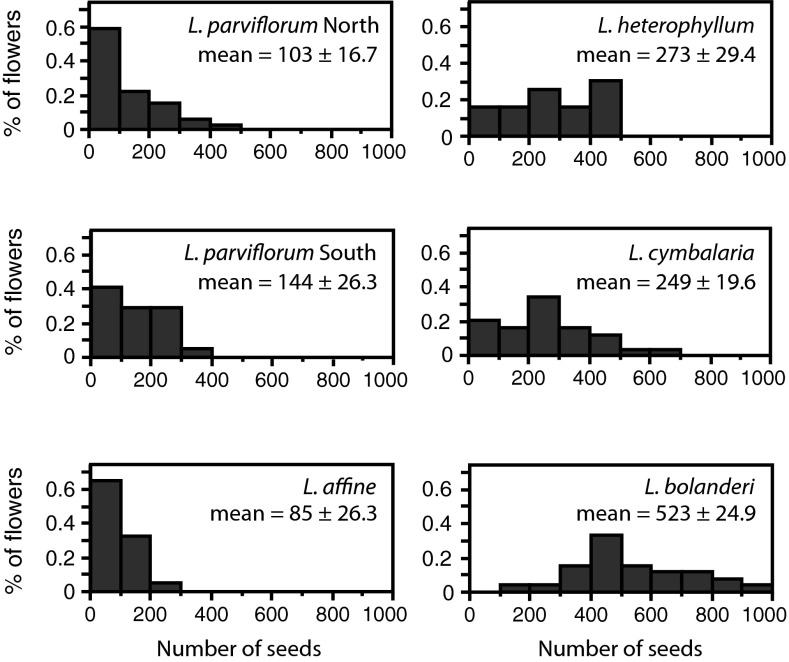
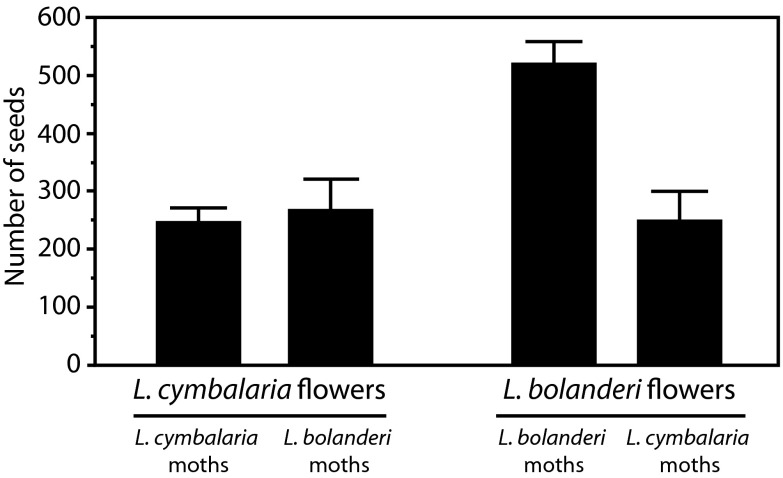
Mutualisms between species are interactions in which reciprocal exploitation results in outcomes that are mutually beneficial. This reciprocal exploitation is evident in the more than a thousand plant species that are pollinated exclusively by insects specialized to lay their eggs in the flowers they pollinate. By pollinating each flower in which she lays eggs, an insect guarantees that her larval offspring have developing seeds on which to feed, whereas the plant gains a specialized pollinator at the cost of some seeds. These mutualisms are often reciprocally obligate, potentially driving not only ongoing coadaptation but also diversification. The lack of known intermediate stages in most of these mutualisms, however, makes it difficult to understand whether these interactions could have begun to diversify even before they became reciprocally obligate. Experimental studies of the incompletely obligate interactions between woodland star (Lithophragma; Saxifragaceae) plants and their pollinating floral parasites in the moth genus Greya (Prodoxidae) show that, as these lineages have diversified, the moths and plants have evolved in ways that maintain effective oviposition and pollination. Experimental assessment of pollination in divergent species and quantitative evaluation of time-lapse photographic sequences of pollination viewed on surgically manipulated flowers show that various combinations of traits are possible for maintaining the mutualism. The results suggest that at least some forms of mutualism can persist and even diversify when the interaction is not reciprocally obligate.
Keywords: coevolution; correlated traits; geographic divergence; trait matching.
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Bascompte J, Jordano P. Plant-animal mutualistic networks: The architecture of biodiversity. Annu Rev Ecol Evol Syst. 2007;38:567–593.
-
- Jordano P, Bascompte J, Olesen JM. Invariant properties in coevolutionary networks of plant-animal interactions. Ecol Lett. 2003;6(1):69–81.
-
- Thompson JN. The Geographic Mosaic of Coevolution. Chicago: Univ of Chicago Press; 2005.
-
- Reynolds RJ, Kula AAR, Fenster CB, Dudash MR. Variable nursery pollinator importance and its effect on plant reproductive success. Oecologia. 2012;168(2):439–448. - PubMed
-
- Godsoe W, Yoder JB, Smith CI, Pellmyr O. Coevolution and divergence in the Joshua tree/yucca moth mutualism. Am Nat. 2008;171(6):816–823. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
