

Mitochondria and FOXO3: breath or die
- PMID: 23801966
- PMCID: PMC3687139
- DOI: 10.3389/fphys.2013.00147
Mitochondria and FOXO3: breath or die
Abstract
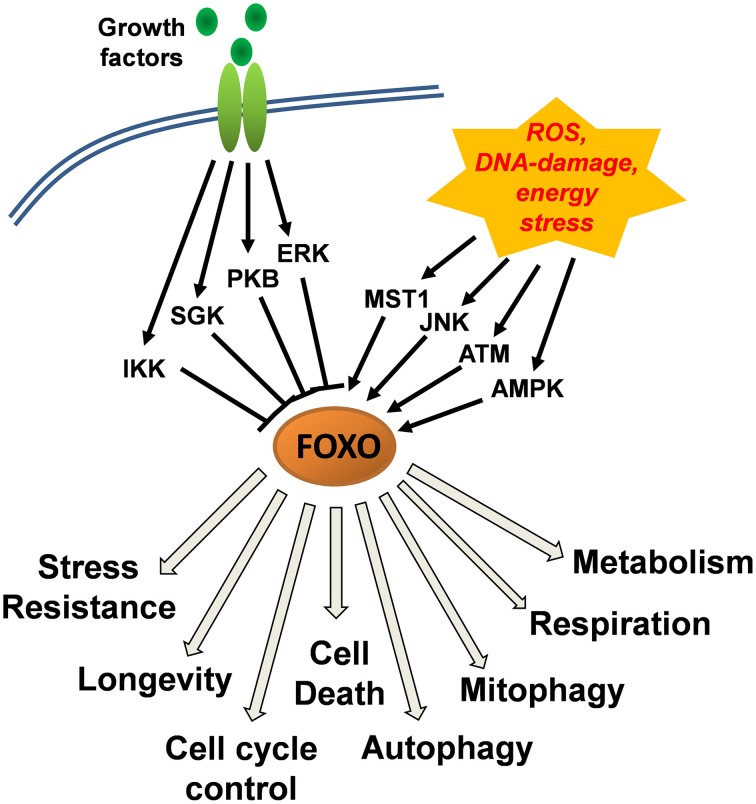
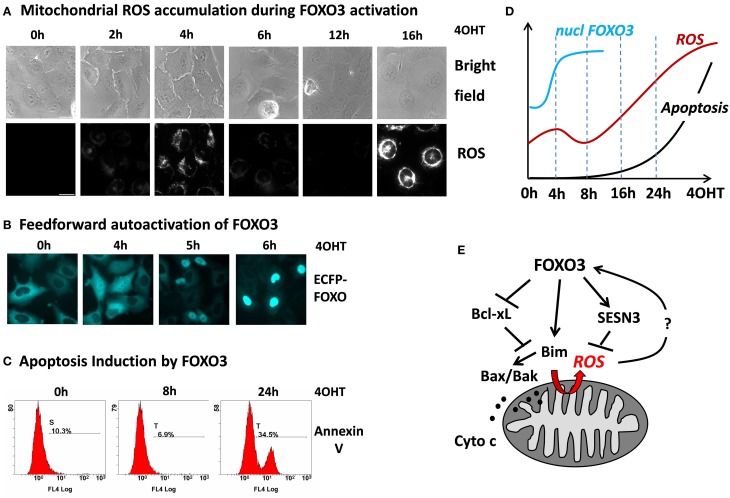
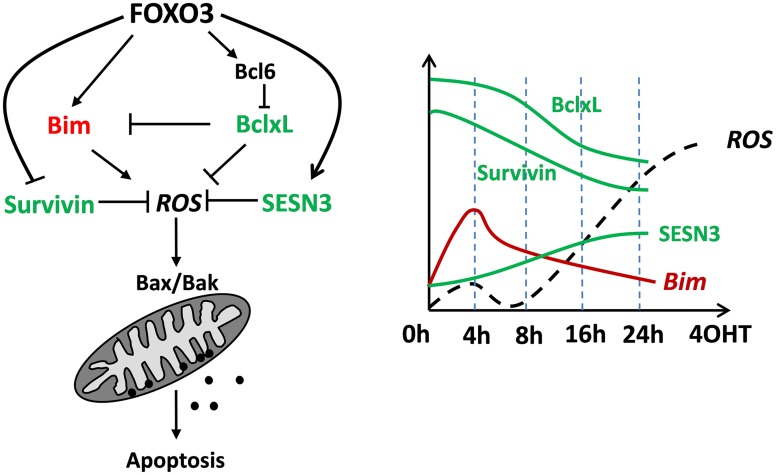
Forkhead box O (FOXO) transcription factors are regulators of cell-type specific apoptosis and cell cycle arrest but also control longevity and reactive oxygen species (ROS). ROS-control by FOXO is mediated by transcriptional activation of detoxifying enzymes such as Superoxide dismutase 2 (SOD2), Catalase or Sestrins or by the repression of mitochondrial respiratory chain proteins resulting in reduced mitochondrial activity. FOXO3 also regulates the adaptation to hypoxia by reducing mitochondrial mass and oxygen consumption during HIF-1α activation. In neuronal tumor cells, FOXO3 triggers ROS-accumulation as a consequence of transient mitochondrial outer membrane permeabilization, which is essential for FOXO3-induced apoptosis in these cells. Cellular ROS levels are affected by the FOXO-targets Bim, BclxL, and Survivin. All three proteins localize to mitochondria and affect mitochondrial membrane potential, respiration and cellular ROS levels. Bim-activation by FOXO3 causes mitochondrial depolarization resulting in a transitory decrease of respiration and ROS production. Survivin, on the other hand, actively changes mitochondrial architecture, respiration-efficacy and energy metabolism. This ability distinguishes Survivin from other anti-apoptotic proteins such as BclxL, which inhibits ROS by inactivating Bim but does not alter mitochondrial function. Importantly, FOXO3 simultaneously also activates ROS-detoxification via induction of SESN3. In this paper we discuss the hypothesis that the delicate balance between ROS-accumulation by Bim-triggered mitochondrial damage, mitochondrial architecture and ROS-detoxifying proteins determines cell fate. We provide evidence for a FOXO self-reactivating loop and for novel functions of FOXO3 in controlling mitochondrial respiration of neuronal cells, which further supports the current view that FOXO transcription factors are information-integrating sentinels of cellular stress and critical modulators of cell homeostasis.
Keywords: BH3-only proteins; Bcl2-rheostat; forkhead transcription factor; mitochondrial fission; mitochondrial respiration.
Figures

References
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials