

Inter-residue coupling contributes to high-affinity subtype-selective binding of α-bungarotoxin to nicotinic receptors
- PMID: 23802200
- PMCID: PMC3912756
- DOI: 10.1042/BJ20130638
Inter-residue coupling contributes to high-affinity subtype-selective binding of α-bungarotoxin to nicotinic receptors
Abstract
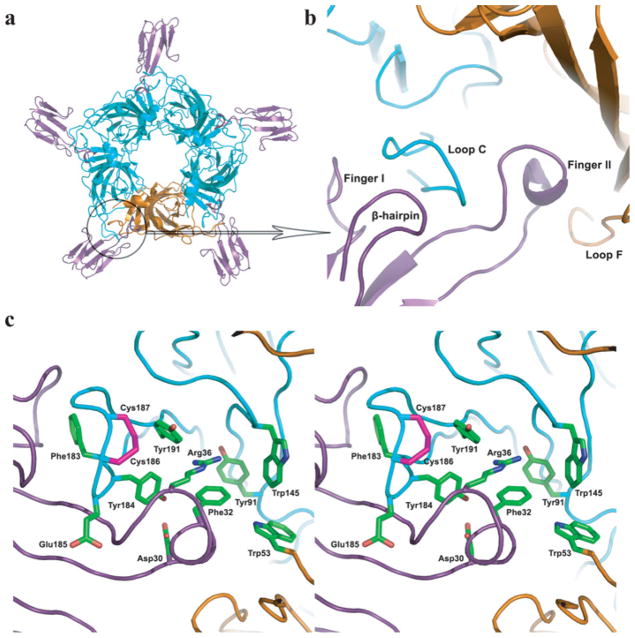
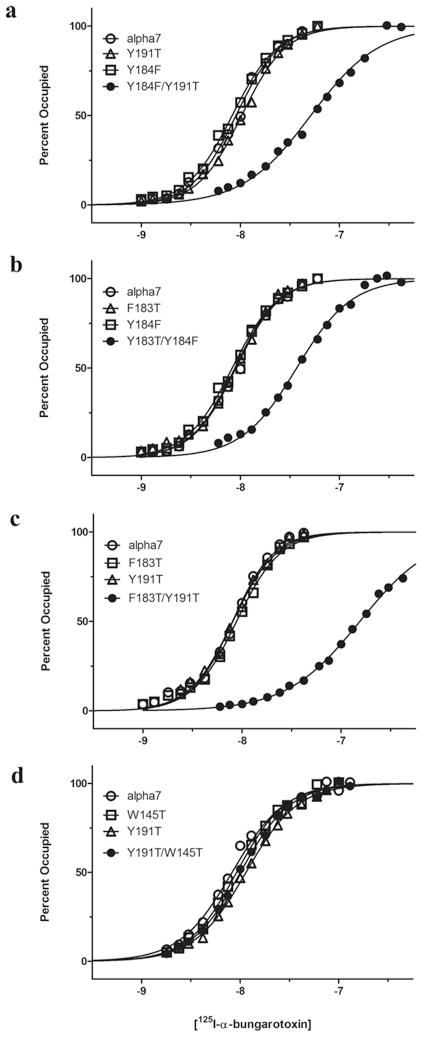
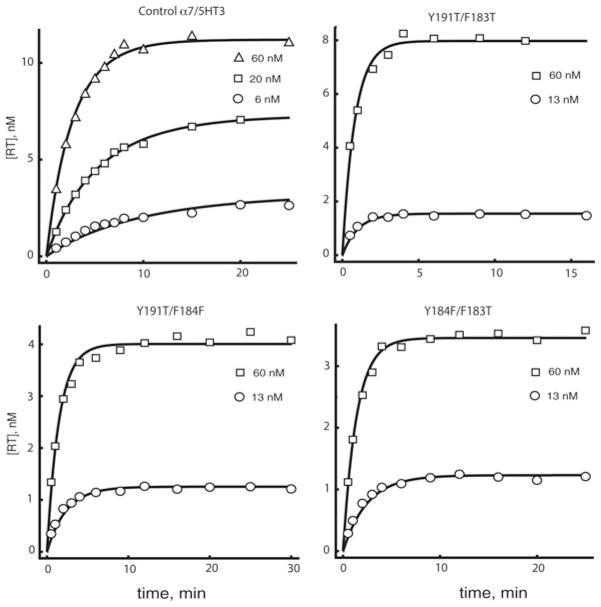
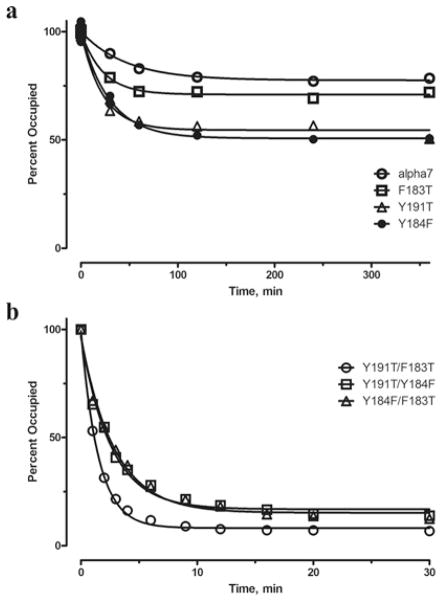
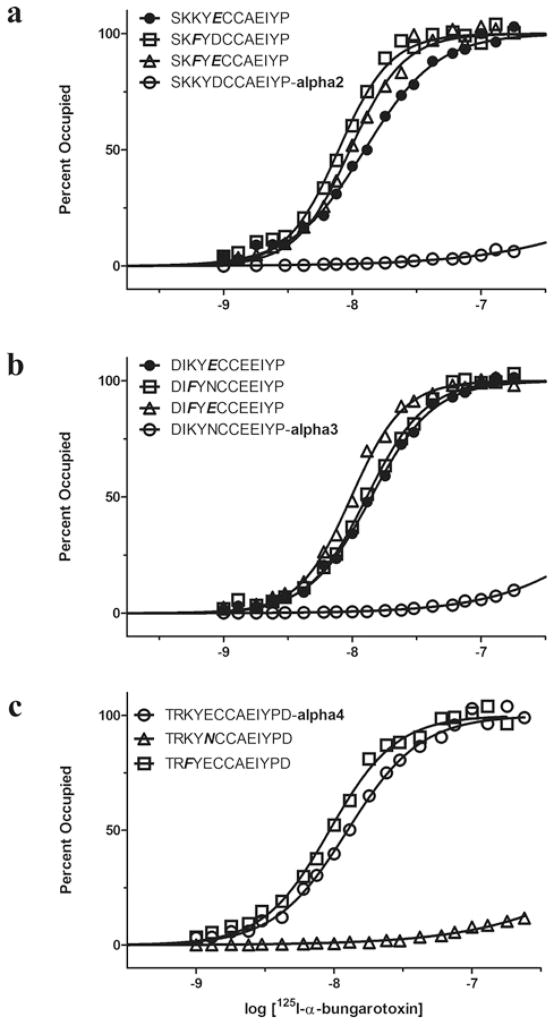
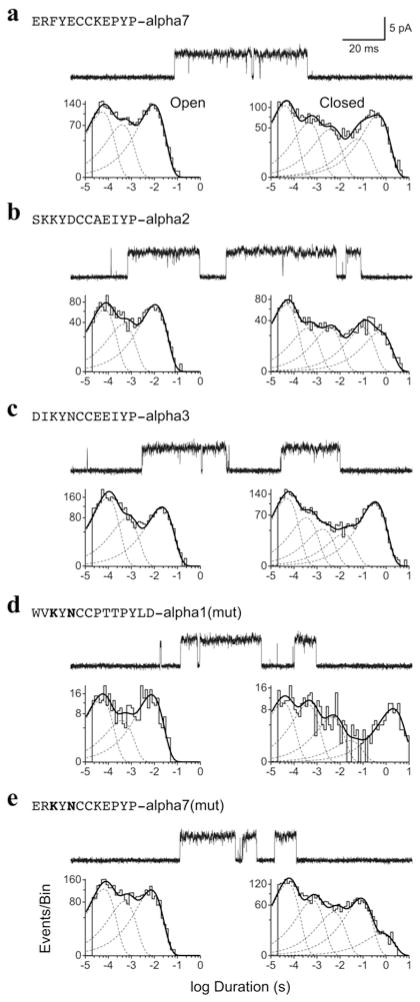
The crystal structure of a pentameric α7 ligand-binding domain chimaera with bound α-btx (α-bungarotoxin) showed that of the five conserved aromatic residues in α7, only Tyr¹⁸⁴ in loop C of the ligand-binding site was required for high-affinity binding. To determine whether the contribution of Tyr¹⁸⁴ depends on local residues, we generated mutations in an α7/5HT(3A) (5-hydroxytryptamine type 3A) receptor chimaera, individually and in pairs, and measured ¹²⁵I-labelled α-btx binding. The results show that mutations of individual residues near Tyr¹⁸⁴ do not affect α-btx affinity, but pairwise mutations decrease affinity in an energetically coupled manner. Kinetic measurements show that the affinity decreases arise through increases in the α-btx dissociation rate with little change in the association rate. Replacing loop C in α7 with loop C from the α-btx-insensitive α2 or α3 subunits abolishes high-affinity α-btx binding, but preserves acetylcholine-elicited single channel currents. However, in both the α2 and α3 construct, mutating either residue that flanks Tyr¹⁸⁴ to its α7 counterpart restores high-affinity α-btx binding. Analogously, in α7, mutating both residues that flank Tyr¹⁸⁴ to the α2 or α3 counterparts abolishes high-affinity α-btx binding. Thus interaction between Tyr¹⁸⁴ and local residues contributes to high-affinity subtype-selective α-btx binding.
Figures


Similar articles
-
Complex between α-bungarotoxin and an α7 nicotinic receptor ligand-binding domain chimaera.Biochem J. 2013 Sep 1;454(2):303-310. doi: 10.1042/BJ20130636. Biochem J. 2013. PMID: 23800261 Free PMC article.
-
Molecular characterization of Dalpha6 and Dalpha7 nicotinic acetylcholine receptor subunits from Drosophila: formation of a high-affinity alpha-bungarotoxin binding site revealed by expression of subunit chimeras.J Neurochem. 2004 Jul;90(2):479-89. doi: 10.1111/j.1471-4159.2004.02499.x. J Neurochem. 2004. PMID: 15228604
-
Water-soluble LYNX1 residues important for interaction with muscle-type and/or neuronal nicotinic receptors.J Biol Chem. 2013 May 31;288(22):15888-99. doi: 10.1074/jbc.M112.436576. Epub 2013 Apr 12. J Biol Chem. 2013. PMID: 23585571 Free PMC article.
-
Localization of agonist and competitive antagonist binding sites on nicotinic acetylcholine receptors.Neurochem Int. 2000 Jun;36(7):595-645. doi: 10.1016/s0197-0186(99)00154-0. Neurochem Int. 2000. PMID: 10771117 Review.
-
Topology of ligand binding sites on the nicotinic acetylcholine receptor.Brain Res Brain Res Rev. 1997 Oct;25(2):133-91. doi: 10.1016/s0165-0173(97)00020-9. Brain Res Brain Res Rev. 1997. PMID: 9403137 Review.
Cited by
-
Structural basis for cooperative interactions of substituted 2-aminopyrimidines with the acetylcholine binding protein.Proc Natl Acad Sci U S A. 2014 Jul 22;111(29):10749-54. doi: 10.1073/pnas.1410992111. Epub 2014 Jul 8. Proc Natl Acad Sci U S A. 2014. PMID: 25006260 Free PMC article.
-
Molecular function of α7 nicotinic receptors as drug targets.J Physiol. 2018 May 15;596(10):1847-1861. doi: 10.1113/JP275101. Epub 2017 Nov 29. J Physiol. 2018. PMID: 29131336 Free PMC article. Review.
-
Receptor variability-driven evolution of snake toxins.Zool Res. 2018 Nov 18;39(6):431-436. doi: 10.24272/j.issn.2095-8137.2018.063. Zool Res. 2018. PMID: 30084433 Free PMC article.
-
Inhibition of nicotinic acetylcholine receptors, a novel facet in the pleiotropic activities of snake venom phospholipases A2.PLoS One. 2014 Dec 18;9(12):e115428. doi: 10.1371/journal.pone.0115428. eCollection 2014. PLoS One. 2014. PMID: 25522251 Free PMC article.
-
Differential Contribution of Subunit Interfaces to α9α10 Nicotinic Acetylcholine Receptor Function.Mol Pharmacol. 2017 Mar;91(3):250-262. doi: 10.1124/mol.116.107482. Epub 2017 Jan 9. Mol Pharmacol. 2017. PMID: 28069778 Free PMC article.
References
-
- Chang CC, Lee CY. Isolation of neurotoxins from the venom of Bungaris multicinctus and their modes of neuromuscular blocking action. Arch Int Pharmacodyn Ther. 1963;144:241–257. - PubMed
-
- Lee CY. Chemistry and pharmacology of polypeptide toxins in snake venoms. Annu Rev Pharmacol. 1972;12:265–286. - PubMed
-
- Weber M, Changeux JP. Binding of Naja nicricollis (3H) α-toxin to membrane fragments from Electropherus and Torpedo electric organs. I Binding of the tritiated α-neurotoxin in the absence of effector. Mol Pharmacol. 1974;10:1–14. - PubMed
-
- Weiland G, Georgia B, Wee VT, Chignell CF, Taylor P. Ligand interactions with cholinergic receptor-enriched membranes from Torpedo: influence of agonist exposure on receptor properties. Mol Pharmacol. 1976;12:1091–1105. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources