

Concurrent maturation of inner hair cell synaptic Ca2+ influx and auditory nerve spontaneous activity around hearing onset in mice
- PMID: 23804089
- PMCID: PMC6618488
- DOI: 10.1523/JNEUROSCI.1215-13.2013
Concurrent maturation of inner hair cell synaptic Ca2+ influx and auditory nerve spontaneous activity around hearing onset in mice
Abstract
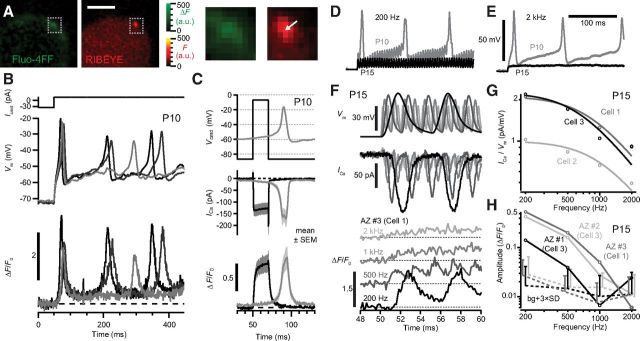
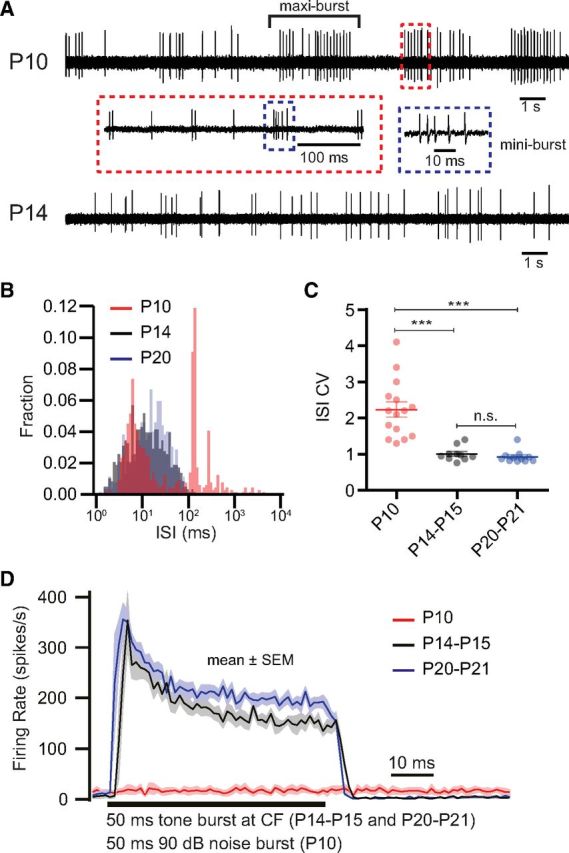
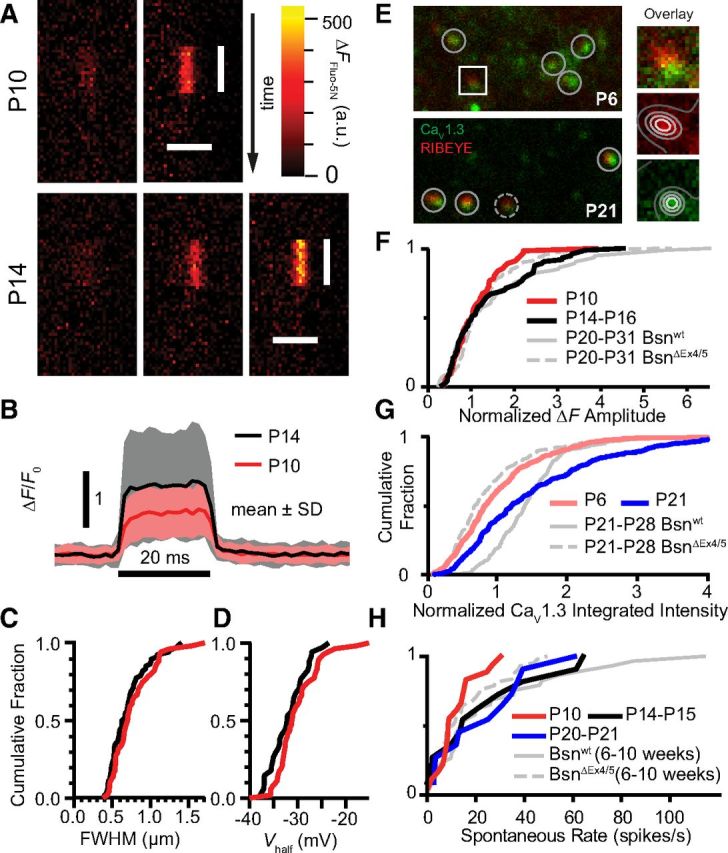
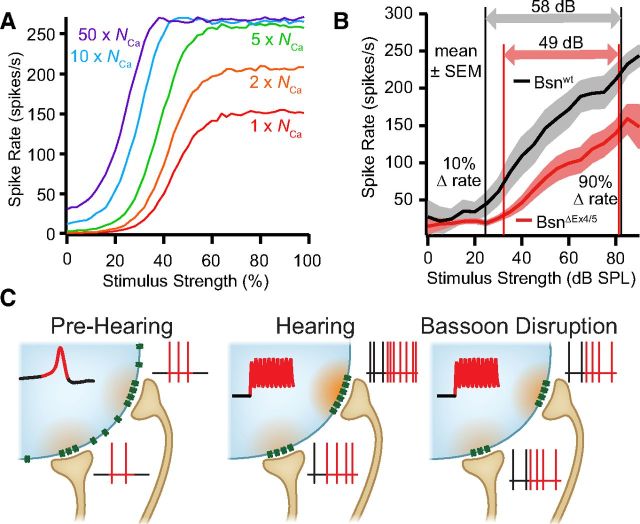
Hearing over a wide range of sound intensities is thought to require complementary coding by functionally diverse spiral ganglion neurons (SGNs), each changing activity only over a subrange. The foundations of SGN diversity are not well understood but likely include differences among their inputs: the presynaptic active zones (AZs) of inner hair cells (IHCs). Here we studied one candidate mechanism for causing SGN diversity-heterogeneity of Ca(2+) influx among the AZs of IHCs-during postnatal development of the mouse cochlea. Ca(2+) imaging revealed a change from regenerative to graded synaptic Ca(2+) signaling after the onset of hearing, when in vivo SGN spike timing changed from patterned to Poissonian. Furthermore, we detected the concurrent emergence of stronger synaptic Ca(2+) signals in IHCs and higher spontaneous spike rates in SGNs. The strengthening of Ca(2+) signaling at a subset of AZs primarily reflected a gain of Ca(2+) channels. We hypothesize that the number of Ca(2+) channels at each IHC AZ critically determines the firing properties of its corresponding SGN and propose that AZ heterogeneity enables IHCs to decompose auditory information into functionally diverse SGNs.
Figures
Similar articles
-
Hair cells use active zones with different voltage dependence of Ca2+ influx to decompose sounds into complementary neural codes.Proc Natl Acad Sci U S A. 2016 Aug 9;113(32):E4716-25. doi: 10.1073/pnas.1605737113. Epub 2016 Jul 26. Proc Natl Acad Sci U S A. 2016. PMID: 27462107 Free PMC article.
-
Bridging the gap between presynaptic hair cell function and neural sound encoding.Elife. 2024 Dec 24;12:RP93749. doi: 10.7554/eLife.93749. Elife. 2024. PMID: 39718472 Free PMC article.
-
Pou4f1 Defines a Subgroup of Type I Spiral Ganglion Neurons and Is Necessary for Normal Inner Hair Cell Presynaptic Ca2+ Signaling.J Neurosci. 2019 Jul 3;39(27):5284-5298. doi: 10.1523/JNEUROSCI.2728-18.2019. Epub 2019 May 13. J Neurosci. 2019. PMID: 31085606 Free PMC article.
-
The afferent signaling complex: Regulation of type I spiral ganglion neuron responses in the auditory periphery.Hear Res. 2016 Jun;336:1-16. doi: 10.1016/j.heares.2016.03.011. Epub 2016 Mar 25. Hear Res. 2016. PMID: 27018296 Review.
-
Diversity matters - extending sound intensity coding by inner hair cells via heterogeneous synapses.EMBO J. 2023 Dec 1;42(23):e114587. doi: 10.15252/embj.2023114587. Epub 2023 Oct 6. EMBO J. 2023. PMID: 37800695 Free PMC article. Review.
Cited by
-
Rab3-interacting molecules 2α and 2β promote the abundance of voltage-gated CaV1.3 Ca2+ channels at hair cell active zones.Proc Natl Acad Sci U S A. 2015 Jun 16;112(24):E3141-9. doi: 10.1073/pnas.1417207112. Epub 2015 Jun 1. Proc Natl Acad Sci U S A. 2015. PMID: 26034270 Free PMC article.
-
EF-hand protein Ca2+ buffers regulate Ca2+ influx and exocytosis in sensory hair cells.Proc Natl Acad Sci U S A. 2015 Mar 3;112(9):E1028-37. doi: 10.1073/pnas.1416424112. Epub 2015 Feb 17. Proc Natl Acad Sci U S A. 2015. PMID: 25691754 Free PMC article.
-
Synaptic Contributions to Cochlear Outer Hair Cell Ca2+ Dynamics.J Neurosci. 2021 Aug 11;41(32):6812-6821. doi: 10.1523/JNEUROSCI.3008-20.2021. Epub 2021 Jul 12. J Neurosci. 2021. PMID: 34253627 Free PMC article.
-
RIM-Binding Protein 2 Promotes a Large Number of CaV1.3 Ca2+-Channels and Contributes to Fast Synaptic Vesicle Replenishment at Hair Cell Active Zones.Front Cell Neurosci. 2017 Nov 2;11:334. doi: 10.3389/fncel.2017.00334. eCollection 2017. Front Cell Neurosci. 2017. PMID: 29163046 Free PMC article.
-
A sensory cell diversifies its output by varying Ca2+ influx-release coupling among active zones.EMBO J. 2021 Mar 1;40(5):e106010. doi: 10.15252/embj.2020106010. Epub 2020 Dec 21. EMBO J. 2021. PMID: 33346936 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous