

Differential impact of a complex environment on positive affect in an animal model of individual differences in emotionality
- PMID: 23806722
- PMCID: PMC3841231
- DOI: 10.1016/j.neuroscience.2013.06.015
Differential impact of a complex environment on positive affect in an animal model of individual differences in emotionality
Abstract
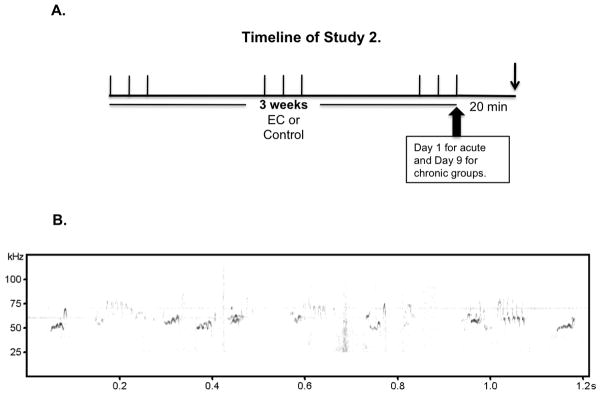
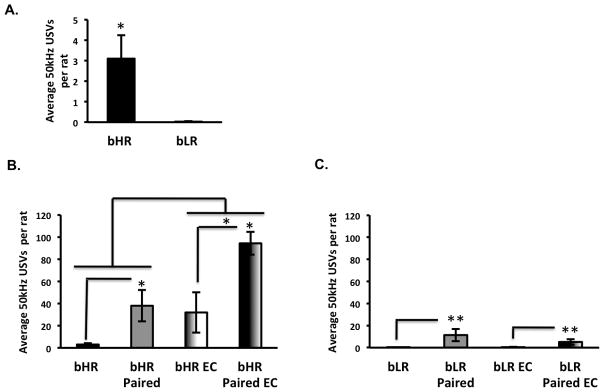
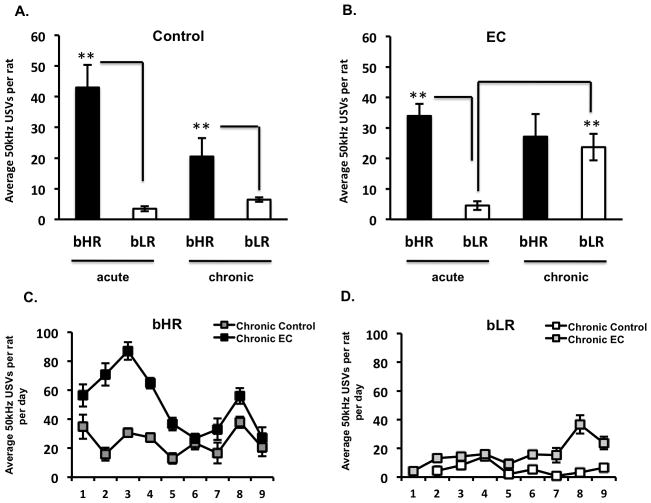
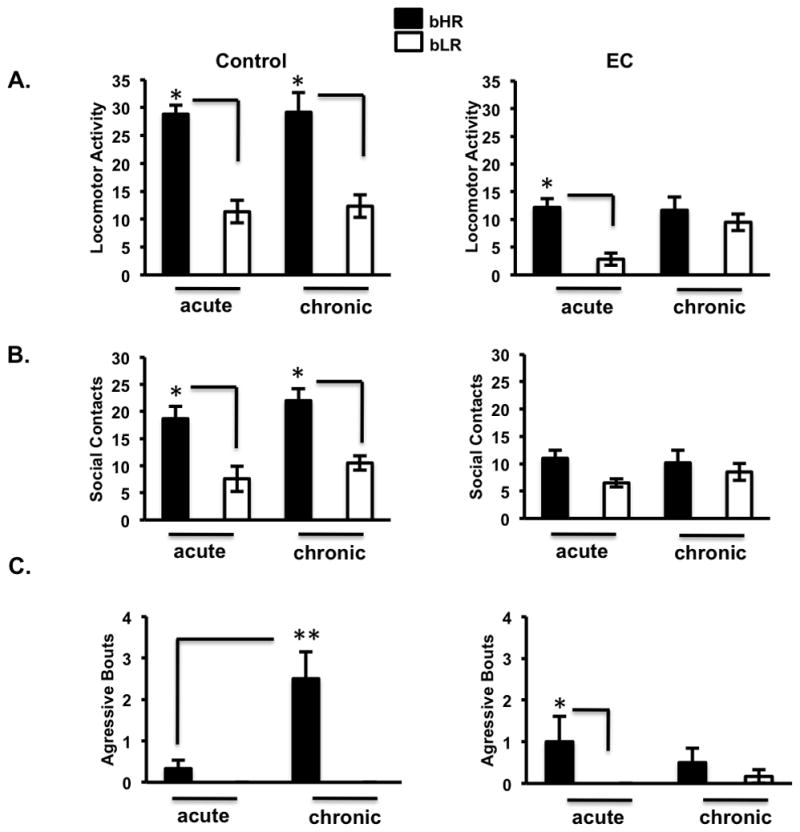
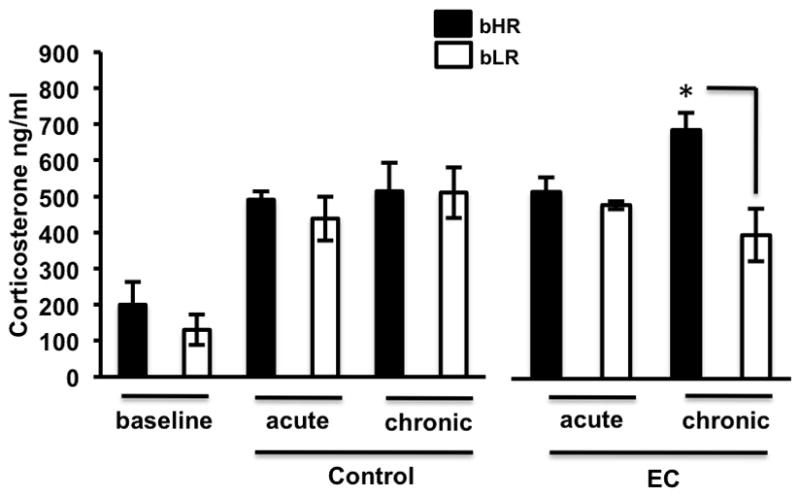
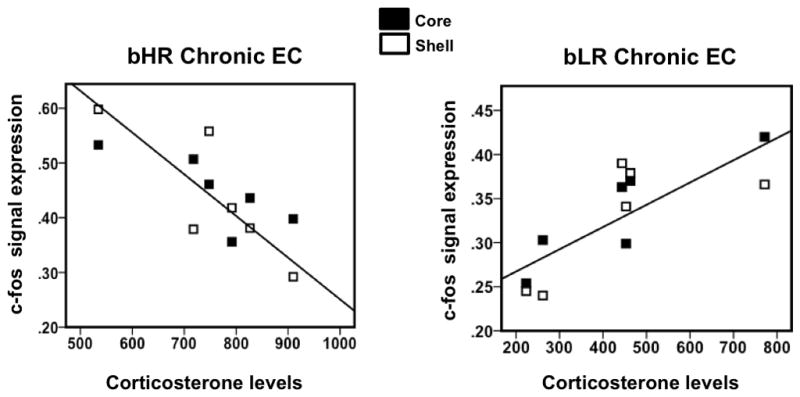
Anhedonia, or the inability to experience positive feelings is a hallmark of depression. However, few animal models have relied on decreased positive affect as an index of susceptibility to depression. Rats emit frequency-modulated ultrasonic vocalizations (USVs), designated as "positive" calls in the 50-kHz range. USVs have been associated with pharmacological activation of motivational reward circuits. Here we utilized selectively-bred rats differing in "emotionality" to ask whether there are associated differences in USVs. Rats bred based on locomotor response to novelty and classified as bred High Responders (bHRs) or bred Low Responders (bLRs) exhibit inborn differences in response to environmental cues, stress responsiveness, and depression-like behavior. These animals also exhibit differences in anxiety-like behavior, which are reversed by exposure to environmental complexity (EC). Finally, these animals exhibit unique profiles of responsiveness to rewarding stimuli accompanied with distinct patterns of dopamine regulation. We investigated whether acute and chronic environmental manipulations impacted USVs in bHRs and bLRs. We found that, relative to bLRs, bHRs emitted significantly more 50-kHz USVs. However, if a bLR is accompanied by another bLR, there is a significant increase in 50-kHZ USVs emitted by this phenotype. bHRs emitted increases in 50-kHZ UVSs upon first exposure to EC, whereas bLRs showed a similar increase only after repeated exposure. bLRs' increase in positive affect after chronic EC was coupled with significant positive correlations between corticosterone levels and c-fos mRNA in the accumbens. Conversely, a decline in the rate of positive calls in bHRs after chronic EC was associated with a negative correlation between corticosterone and accumbens c-fos mRNA. These studies demonstrate that inborn differences in emotionality interact with the environment to influence positive affect and underscore the potential interaction between glucocorticoids and the mesolimbic reward circuitry in modulating 50-kHz calls.
Keywords: affect; emotionality; individual differences; ultrasonic vocalization.
Published by Elsevier Ltd.
Figures
Similar articles
-
Effects of early-life FGF2 on ultrasonic vocalizations (USVs) and the mu-opioid receptor in male Sprague-Dawley rats selectively-bred for differences in their response to novelty.Brain Res. 2019 Jul 15;1715:106-114. doi: 10.1016/j.brainres.2019.03.011. Epub 2019 Mar 14. Brain Res. 2019. PMID: 30880118 Free PMC article.
-
An animal model of genetic vulnerability to behavioral disinhibition and responsiveness to reward-related cues: implications for addiction.Neuropsychopharmacology. 2010 Jan;35(2):388-400. doi: 10.1038/npp.2009.142. Neuropsychopharmacology. 2010. PMID: 19794408 Free PMC article.
-
High novelty-seeking rats are resilient to negative physiological effects of the early life stress.Stress. 2014 Jan;17(1):97-107. doi: 10.3109/10253890.2013.850670. Epub 2013 Nov 6. Stress. 2014. PMID: 24090131 Free PMC article.
-
Rat 50-kHz ultrasonic vocalizations as a tool in studying neurochemical mechanisms that regulate positive emotional states.J Neurosci Methods. 2018 Dec 1;310:33-44. doi: 10.1016/j.jneumeth.2018.06.018. Epub 2018 Jun 26. J Neurosci Methods. 2018. PMID: 29959002 Review.
-
Frequency-modulated 50 kHz ultrasonic vocalizations: a tool for uncovering the molecular substrates of positive affect.Neurosci Biobehav Rev. 2011 Oct;35(9):1831-6. doi: 10.1016/j.neubiorev.2010.11.011. Epub 2010 Dec 7. Neurosci Biobehav Rev. 2011. PMID: 21144859 Review.
Cited by
-
Treatment resistant depression: A multi-scale, systems biology approach.Neurosci Biobehav Rev. 2018 Jan;84:272-288. doi: 10.1016/j.neubiorev.2017.08.019. Epub 2017 Aug 30. Neurosci Biobehav Rev. 2018. PMID: 28859997 Free PMC article. Review.
-
Applying evidence-based cross-disciplinary concepts helps to explain the heterogeneity in pain, function, and biological measures in individuals with knee pain with/at risk of osteoarthritis.Pain Rep. 2024 Dec 24;10(1):e1225. doi: 10.1097/PR9.0000000000001225. eCollection 2025 Feb. Pain Rep. 2024. PMID: 39726853 Free PMC article.
-
Conflict between Threat Sensitivity and Sensation Seeking in the Adolescent Brain: Role of the Hippocampus, and Neurobehavioural Plasticity Induced by Pleasurable Early Enriched Experience.Brain Sci. 2021 Feb 20;11(2):268. doi: 10.3390/brainsci11020268. Brain Sci. 2021. PMID: 33672653 Free PMC article.
-
Adolescent environmental enrichment induces social resilience and alters neural gene expression in a selectively bred rodent model with anxious phenotype.Neurobiol Stress. 2024 May 30;31:100651. doi: 10.1016/j.ynstr.2024.100651. eCollection 2024 Jul. Neurobiol Stress. 2024. PMID: 38933284 Free PMC article.
-
Positive Emotional Learning Induces Resilience to Depression: A Role for NMDA Receptor-mediated Synaptic Plasticity.Curr Neuropharmacol. 2017;15(1):3-10. doi: 10.2174/1570159x14666160422110344. Curr Neuropharmacol. 2017. PMID: 27102428 Free PMC article. Review.
References
-
- Burgdorf J, Knutson B, Panksepp J. Anticipation of rewarding electrical brain stimulation evokes ultrasonic vocalization in rats. Behav Neurosci. 2000;114:320–327. - PubMed
-
- Burgdorf J, Knutson B, Panksepp J, Ikemoto S. Nucleus accumbens amphetamine microinjections unconditionally elicit 50-kHz ultrasonic vocalizations in rats. Behav Neurosci. 2001;115:940–944. - PubMed
-
- Burgdorf J, Panksepp J. The neurobiology of positive emotions. Neurosci Biobehav Rev. 2006;30:173–187. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
