

Epigenetic pathways and glioblastoma treatment
- PMID: 23807265
- PMCID: PMC3883781
- DOI: 10.4161/epi.25440
Epigenetic pathways and glioblastoma treatment
Abstract
Glioblastoma multiforme (GBM) is the most common malignant adult brain tumor. Standard GBM treatment includes maximal safe surgical resection with combination radiotherapy and adjuvant temozolomide (TMZ) chemotherapy. Alarmingly, patient survival at five-years is below 10%. This is in part due to the invasive behavior of the tumor and the resulting inability to resect greater than 98% of some tumors. In fact, recurrence after such treatment may be inevitable, even in cases where gross total resection is achieved. The Cancer Genome Atlas (TCGA) research network performed whole genome sequencing of GBM tumors and found that GBM recurrence is linked to epigenetic mechanisms and pathways. Central to these pathways are epigenetic enzymes, which have recently emerged as possible new drug targets for multiple cancers, including GBM. Here we review GBM treatment, and provide a systems approach to identifying epigenetic drivers of GBM tumor progression based on temporal modeling of putative GBM cells of origin. We also discuss advances in defining epigenetic mechanisms controlling GBM initiation and recurrence and the drug discovery considerations associated with targeting epigenetic enzymes for GBM treatment.
Keywords: drug discovery; epigenetics; glioblastoma; statistical modeling.
Figures
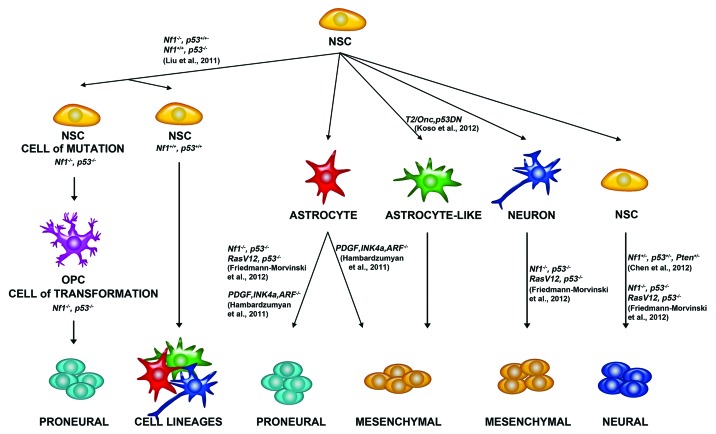
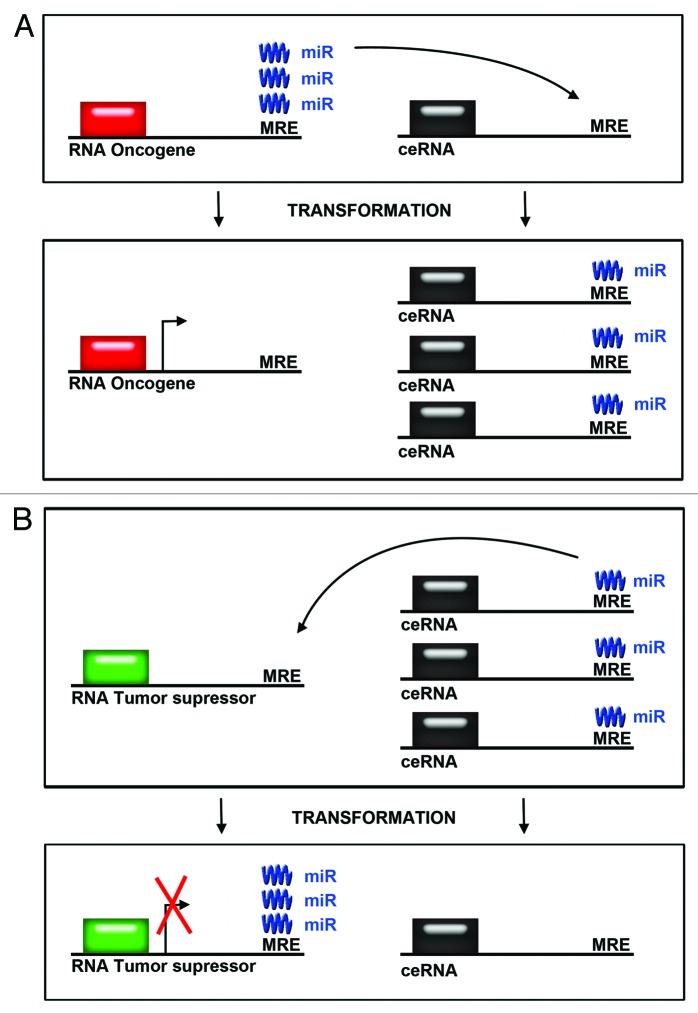
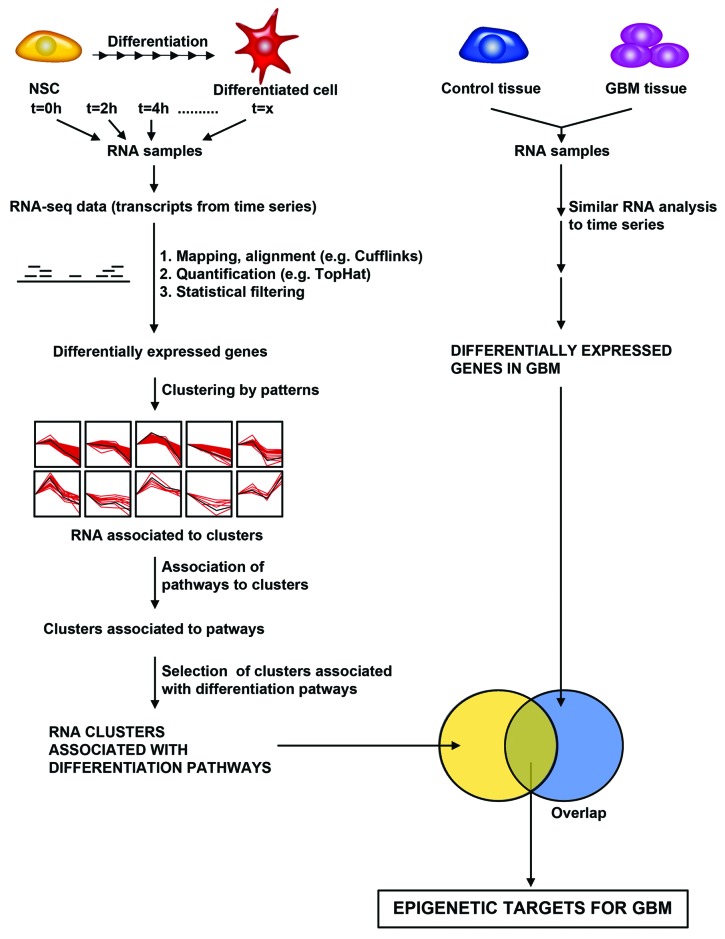
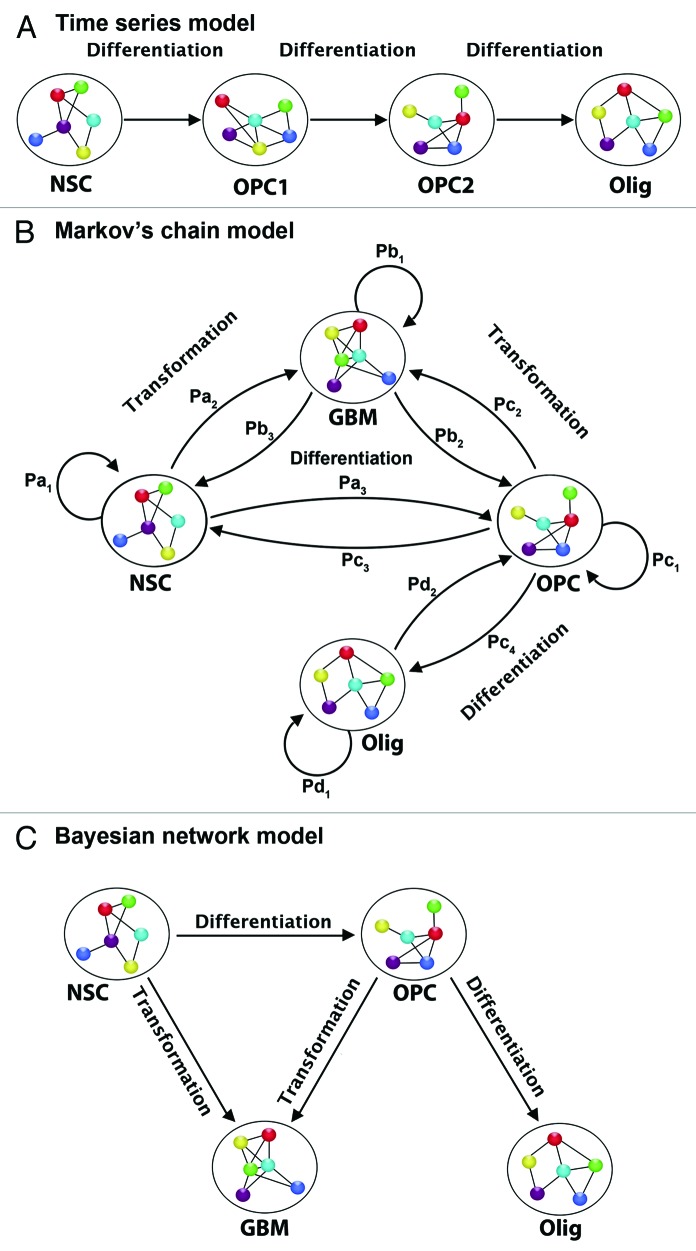
Similar articles
-
The value of temozolomide in combination with radiotherapy during standard treatment for newly diagnosed glioblastoma.J Neurooncol. 2013 Apr;112(2):277-83. doi: 10.1007/s11060-013-1060-3. Epub 2013 Feb 2. J Neurooncol. 2013. PMID: 23377829
-
Phase I study of hypofractionated intensity modulated radiation therapy with concurrent and adjuvant temozolomide in patients with glioblastoma multiforme.Radiat Oncol. 2013 Feb 20;8:38. doi: 10.1186/1748-717X-8-38. Radiat Oncol. 2013. PMID: 23425509 Free PMC article. Clinical Trial.
-
Longitudinal heterogeneity in glioblastoma: moving targets in recurrent versus primary tumors.J Transl Med. 2019 Mar 20;17(1):96. doi: 10.1186/s12967-019-1846-y. J Transl Med. 2019. PMID: 30894200 Free PMC article.
-
An update on the epigenetics of glioblastomas.Epigenomics. 2016 Sep;8(9):1289-305. doi: 10.2217/epi-2016-0040. Epub 2016 Sep 2. Epigenomics. 2016. PMID: 27585647 Review.
-
Problems of Glioblastoma Multiforme Drug Resistance.Biochemistry (Mosc). 2016 Feb;81(2):91-100. doi: 10.1134/S0006297916020036. Biochemistry (Mosc). 2016. PMID: 27260389 Review.
Cited by
-
BPTF regulates growth of adult and pediatric high-grade glioma through the MYC pathway.Oncogene. 2020 Mar;39(11):2305-2327. doi: 10.1038/s41388-019-1125-7. Epub 2019 Dec 16. Oncogene. 2020. PMID: 31844250 Free PMC article.
-
SUZ12 inhibition attenuates cell proliferation of glioblastoma via post-translational regulation of CDKN1B.Genes Genomics. 2023 Dec;45(12):1623-1632. doi: 10.1007/s13258-023-01468-5. Epub 2023 Oct 19. Genes Genomics. 2023. PMID: 37856053
-
Prucalopride inhibits the glioma cells proliferation and induces autophagy via AKT-mTOR pathway.BMC Neurol. 2018 Jun 4;18(1):80. doi: 10.1186/s12883-018-1083-7. BMC Neurol. 2018. PMID: 29866060 Free PMC article.
-
Romo1 is associated with ROS production and cellular growth in human gliomas.J Neurooncol. 2015 Jan;121(1):73-81. doi: 10.1007/s11060-014-1608-x. Epub 2014 Sep 6. J Neurooncol. 2015. PMID: 25193023
-
Chimeric Element-Regulated MRI Reporter System for Mediation of Glioma Theranostics.Cancers (Basel). 2025 Jul 15;17(14):2349. doi: 10.3390/cancers17142349. Cancers (Basel). 2025. PMID: 40723232 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical