

Beyond taxol: microtubule-based treatment of disease and injury of the nervous system
- PMID: 23811322
- PMCID: PMC3784279
- DOI: 10.1093/brain/awt153
Beyond taxol: microtubule-based treatment of disease and injury of the nervous system
Abstract
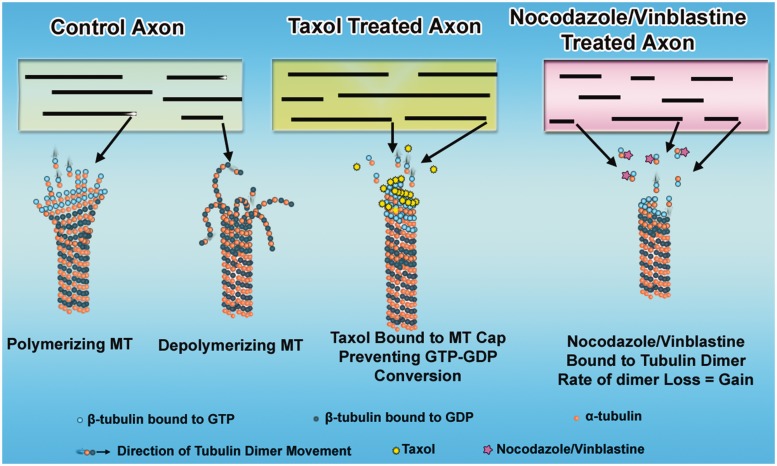
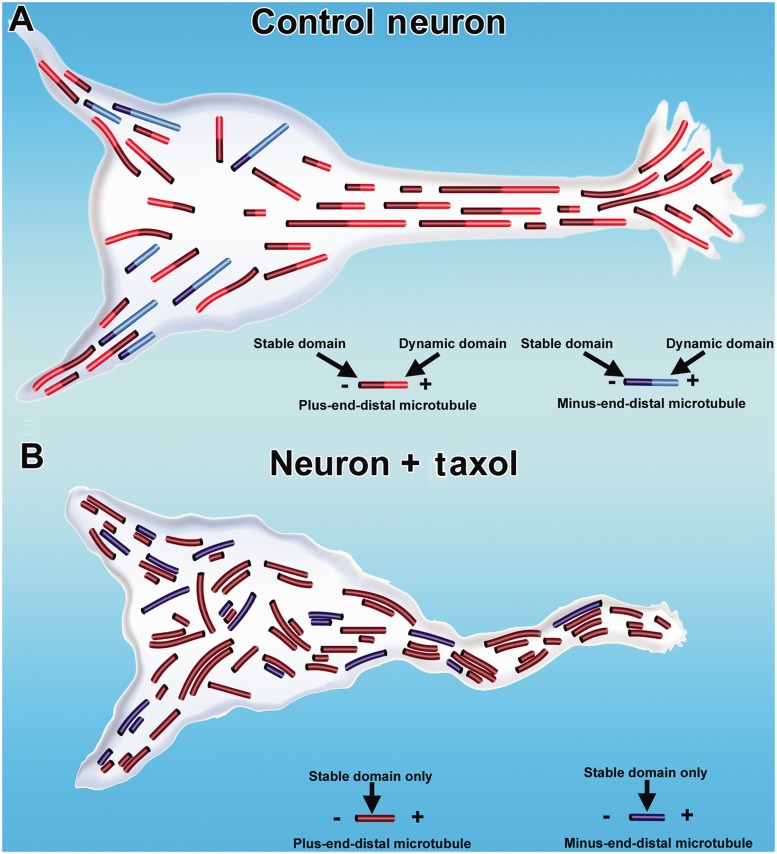
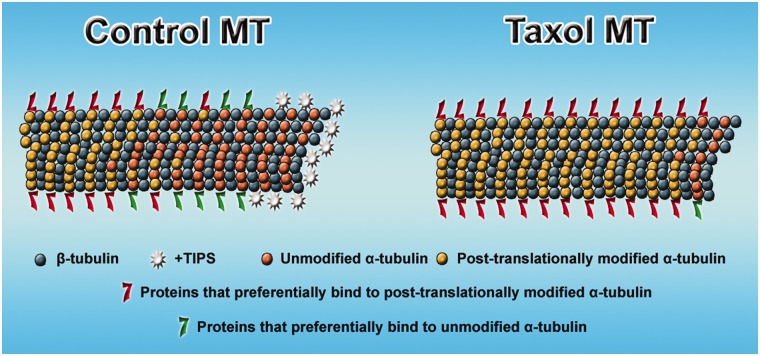
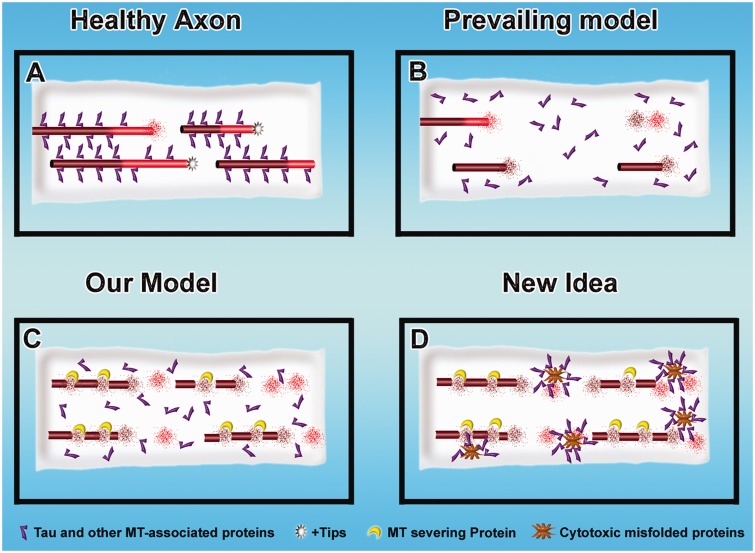
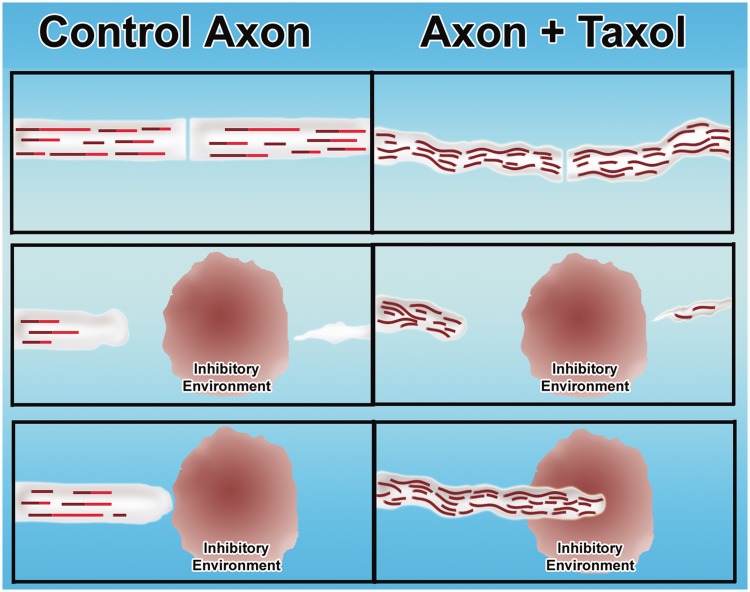
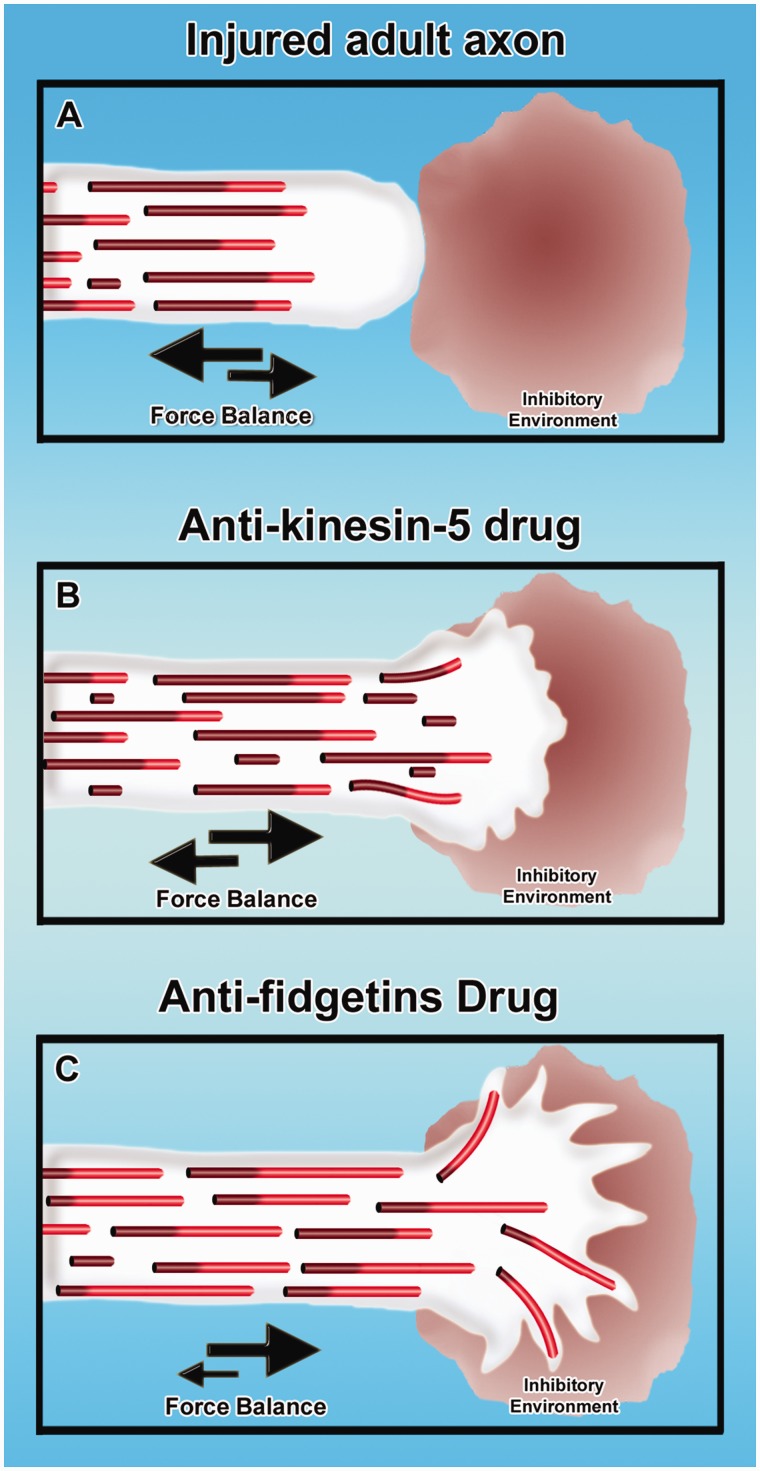
Contemporary research has revealed a great deal of information on the behaviours of microtubules that underlie critical events in the lives of neurons. Microtubules in the neuron undergo dynamic assembly and disassembly, bundling and splaying, severing, and rapid transport as well as integration with other cytoskeletal elements such as actin filaments. These various behaviours are regulated by signalling pathways that affect microtubule-related proteins such as molecular motor proteins and microtubule severing enzymes, as well as a variety of proteins that promote the assembly, stabilization and bundling of microtubules. In recent years, translational neuroscientists have earmarked microtubules as a promising target for therapy of injury and disease of the nervous system. Proof-of-principle has come mainly from studies using taxol and related drugs to pharmacologically stabilize microtubules in animal models of nerve injury and disease. However, concerns persist that the negative consequences of abnormal microtubule stabilization may outweigh the positive effects. Other potential approaches include microtubule-active drugs with somewhat different properties, but also expanding the therapeutic toolkit to include intervention at the level of microtubule regulatory proteins.
Keywords: axon; dendrite; microtubule; neuron; taxol.
Figures
References
-
- Aguirre-Portoles C, Bird AW, Hyman A, Canamero M, Perez de Castro I, Malumbres M. Tpx2 controls spindle integrity, genome stability, and tumor development. Cancer Res. 2012;72:1518–28. - PubMed
-
- Akhmanova A, Steinmetz MO. Tracking the ends: a dynamic protein network controls the fate of microtubule tips. Nat Rev Mol Cell Biol. 2008;9:309–22. - PubMed
-
- Amos LA, Löwe J. How taxol stabilises microtubule structure. Chem Biol. 1999;6:R65–9. - PubMed
-
- Baas PW. Microtubules and neuronal polarity: lessons from mitosis. Neuron. 1999;22:23–31. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
